


Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
СТАТИСТИЧЕСКОЕ ОЦЕНИВАНИЕ РАЗНООБРАЗИЯ РАСТИТЕЛЬНОСТИ НА ОСНОВЕ НЕОДНОРОДНЫХ ГЕОБОТАНИЧЕСКИХ ДАННЫХ
Смирнов В.Э., Ханина Л.Г.
Институт математических проблем биологии РАН, Пущино, Россия vesmirnov@gmail.com
Аннотация: разработана и апробирована методика статистического оценивания параметров видового разнообразия растительности. Проведено сравнение видового разнообразия сосудистых растений для выделенных ранее типов растительных сообществ заповедника «Калужские засеки». Сравнение проведено как для наблюдаемого, так и для эффективного числа видов (чисел Хилла).
Основными оценками разнообразия растительности той или иной территории являются оценки ее экосистемного и видового разнообразия. Экосистемное разнообразие оценивается, как правило, через список типов сообществ, представленных на территории. Оценку видового разнообразия можно проводить на основе геоботанических описаний – списков видов растений на площадках фиксированного размера, с указанием обилия видов в разных ярусах растительного сообщества. В этом случае геоботанические описания классифицируют по типам растительных сообществ, и описания, выполненные в сообществе одного типа, используют для оценки видового разнообразия этого типа.
Видовое разнообразие сообществ оценивают по составляющим их компонентам: альфа разнообразие можно оценить через видовую насыщенность, рассчитываемую как среднее число видов на площадке фиксированного размера; бета разнообразие можно оценить с помощью различных мер сходства/расстояния; локальное гамма разнообразие можно оценить через видовое богатство, которое, как правило, рассчитывают как общее число видов, встреченных на всех геоботанических площадках в сообществе данного типа. При таком подходе оценки видового разнообразия в сильной степени зависят от числа геоботанических описаний, доступных для сообществ. Кроме того, эти оценки очевидно зависят от качества проведенных геоботанических исследований, расположения площадок, их размеров и других параметров. Все эти параметры часто оказываются неоднородными, а количественные оценки разнообразия, полученные на основе таких неоднородных данных, могут оказаться существенно смещенными. Относительно корректные оценки видового разнообразия вместе с оценкой их статистической значимости в условиях неполных или неоднородных геоботанических данных можно получить при использовании специальных статистических методов, активно развиваемых в последние годы (Colwell et al., 2012; Смирнов и др., 2014; Chao et al., 2014; Ханина и др., в печати).
Сравнительные оценки видового богатства в условиях неравномерных выборок описаний мы рассчитывали с помощью видовых кумулятивных кривых в рамках интегрального подхода (Colwell et al., 2012). Для данных по присутствию/отсутствию видов авторы подхода предложили аналитический метод на основе распределения Бернулли, который находит ожидаемое число видов, как в случае интерполяции выборки (приведения выборки большего размера к меньшему), так и в случае ее экстраполяции (доведения выборки меньшего размера до большего). В результате строится единая видовая кумулятивная кривая для каждого типа сообщества, которая начинается от размера выборки в одно описание, продолжается до наблюдаемого числа описаний в данном типе, далее переходит в асимптотическую кривую, продолжающуюся до желаемого уровня экстраполяции. Недостатком этого подхода является то обстоятельство, что надежная экстраполяция видового богатства по данному методу возможна только до удвоенного (максимум утроенного) размера референтной выборки, тогда как число доступных описаний для отдельных типов сообществ может отличаться на порядки. В результате данные для некоторых типов сообществ приходится интерполировать, что, очевидно, ведет к потере
174
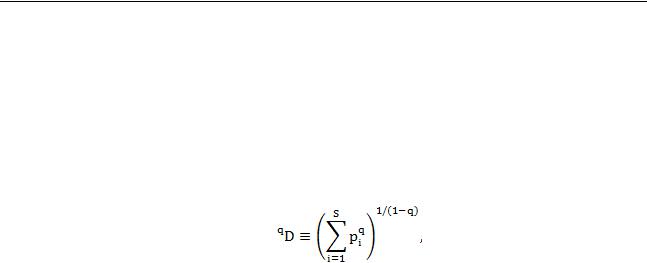
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
информации. Так, например, для сравнительной оценки видового богатства сообществ заповедника «Калужские засеки» 306 геоботанических описаний неморальных широколиственных лесов нам пришлось интерполировать до 57 описаний – числа, равного утроенному размеру минимальной в массиве выборки (Смирнов и др., 2014).
Развитием подхода построения кумулятивных кривых явилось его распространение на такие меры разнообразия, экстраполяция которых возможна до бóльших пределов, нежели экстраполяция видового богатства. Этому требованию, как показали Chao et al. (2014), удовлетворяют некоторые меры разнообразия, входящие в так называемый ряд Хилла (иначе
– числа Хилла). Этот ряд можно представить в виде следующего общего выражения (по Jost,
2006):
где q – константа, определяющая конкретное число Хилла, S – число видов в выборке, pi – относительное обилие i-го вида. Hill (1973) показал, что q = 0 дает число видов в сообществе (видовое богатство), q = 1 соответствует экспоненте индекса Шеннона (в пределе), q = 2 дает величину, обратную индексу Симпсона. Преимущество этих мер в том, что они выражены через число видов, а не являются безразмерными, как оригинальные индексы Шеннона и Симпсона. Рассчитанное таким образом число видов называется эффективным или эквивалентным и соответствует ожидаемому числу видов с равным обилием в анализируемом типе сообщества (Jost, 2006). При этом числа Хилла устроены таким образом, что с увеличением q увеличивается вес (влияние) видов с бóльшим обилием в расчете меры разнообразия. Это позволяет с помощью трех мер (при q = 0, 1, 2) выявлять различные особенности структуры сообщества, релевантные с точки зрения оценки разнообразия (роль редких, часто встречающихся, доминируюших и др. видов). Преимущество эффективных чисел видов состоит также в том, что их значения при экстраполяции быстро выходят на плато и далее изменяются слабо, что увеличивает пределы их практической экстраполяции, в частности до максимального размера выборки в массиве исследуемых данных (Chao et al., 2014). Это было использовано нами для развития методики оценки видового разнообразия и применения ее для оценки видового богатства выделенных ранее типов растительных сообществ заповедника «Калужские засеки».
Полученные нами результаты оценок видового разнообразия сосудистых растений по числам Хилла в целом соответствовали, а в частностях уточняли результаты, полученные ранее (Ханина и др., в печати). Использование чисел Хилла позволило провести более детальный анализ структуры видового разнообразия и учесть весь объем геоботанической информации, доступной для анализа.
Работа проводилась при частичной поддержке РФФИ, проект № 13-04-02181-а.
Литература
Смирнов В.Э., Ханина Л.Г. Бобровский М.В. Оценка видового разнообразия растительности на основе интегрального статистического подхода в условиях неоднородных данных (на примере заповедника «Калужские засеки») // Вестник Томского гос. университета. Биология. - 2014. - № 4 (28). - C. 84–101.
Ханина Л.Г., Смирнов В.Э., Бобровский М.В. Оценка видового богатства растительных сообществ заповедника «Калужские засеки» на основе индексов Шеннона и Симпсона и чисел Хилла // Лесоведение. - В печати.
Chao A., Gotelli N.J., Hsieh T.C., Sander E.L., Ma K.H., Colwell R.K., Ellison A.M. Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies // Ecological monographs. - 2014. - V. 84. - No. 1. - P. 45–67.
Colwell R.K., Chao A., Gotelli N.J., Lin S.-Y., Mao C.X., Chazdon R.L., Longino J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages // Journal of plant ecology. - 2012. - V. 5. - P. 3–21.
Hill M.O. Diversity and evenness: a unifying notation and consequences // Ecology. - 1973. - V. 54. - P. 427–432. Jost L. Entropy and diversity // Oikos. - 2006. - V. 113. - No. 2. - P. 363–375.
175

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ДИНАМИКА ЧИСЛЕННОСТИ ЛЕСНЫХ НАСЕКОМЫХ: ARMA-МОДЕЛИ
Суховольский В.Г. 1, Тарасова О.В.2
1Институт леса им. В.Н. Сукачева СО РАН, Красноярск, Россия soukhovolsky@yandex.ru
2Сибирский федеральный университет, Красноярск, Россия olvitarasova2010@yandex.ru
Аннотация: рассмотрены методы моделирования динамики численности насекомых-филлофагов с использвоанием ARMA–моделей. Использование ARMA-моделей позволило оценить вклад погодных факторов в изменение плотности популяций филлофагов, определить запаздывание реакции паразитов на изменение плотности популяции вида-хозяина.
Два класса факторов – модифицирующие (в частности, погодные) и регулирующие (такие, как воздействия паразитов и хищников) влияют на плотность популяций насекомыхфиллофагов (Исаев и др, 2001). Поэтому для моделирования и прогноза популяционной динамики филлофагов необходимо, кроме данных по плотности популяции моделируемого вида насекомых, знать плотность популяций паразитов и хищников и функции восприимчивости моделируемых популяций насекомых к погодным и ландшафтным факторам. Однако во многих случаях эти переменные неизвестны, что затрудняет построение моделей.
В настоящей работе рассмотрены методы моделирования динамики численности насекомых-филлофагов, позволяющие не включать в модели влияние регулирующих факторов и не определять функцию восприимчивости популяции насекомых к модифицирующим факторам.
Для моделирования и краткосрочного прогноза популяционной динамики насекомых-
филлофагов использованы ARMA (AutoRegression Moving Average) – модели (Бокс,
Дженкинс, 1974; Большаков, Каримов, 2007):
n |
m |
|
L( i + 1 ) = L0 + ∑a j L( i − j ) + ∑bkW ( i − k ) |
(1) |
|
j=1 |
k=0 |
|
где L(i) - регуляризованная нормализованная логарифическая (РНЛ) плотность популяции в сезон i; W(i) – погодный показатель (гидротермический коэффициент вегетационного сезона), L0. aj, bk, n, m – коэффициенты.
В качестве объектов моделирования использовались временные ряды динамики численности серой лиственничной листовертки Zeiraphera griseana L. в Альпах
(Baltensweiler, 1964; Baltensweiler, Fischlin, 1988) сосновой пяденицы Bupalus piniarius L. в
сосновых борах Тюрингии (Германия) (Schwerdtfeger, 1952), сосновой пяденицы Bupalus piniarius L. и соснового шелкопряда Dendrolimus pini L. на юге Средней Сибири (Пальникова и др, 2002; Пальникова и др, 2014), зеленой дубовой листовертки Tortrix viridana L. в европейской части России (Голубев, Багдатьева, 2013).
Модель (1) позволила оценить вклад погодных факторов в изменение плотности популяции, определить запаздывание реакции паразитов на изменение плотности популяции вида-хозяина.
Для ARMA-моделей популяционной динамики лесных насекомых были рассчитаны запас по устойчивости и робастная устойчивость (Гайдук и др, 2011; Ким, 2007; Поляк, Щербаков, 2002). Это позволяет оценить изменение состояния популяции при трансформации внешней среды и изменениях собственных характеристик популяции.
Предложенный подход к моделированию популяционной динамики лесных насекомых-филлофагов позволил получить краткосрочный (на 2-4 года) прогноз динамики численности моделируемых популяций.
176

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
Работа поддержана РФФИ (грант 15-04-01192).
Литература
Бокс Дж., Дженкинс Г. Анализ временных рядов. Прогноз и управление. Вып. 1. М.: Мир, 1974. – 197 с. Большаков А.А., Каримов Р.Н. Методы обработки многомерных данных и временных рядов. М.: Горячая линия
– Телеком, 2007. - 522 с.
Гайдук А.Р., Беляев В.Е., Пьявченко Т.А. Теория автоматического управления в примерах и задачах с решениями в MATLAB. СПб.: Лань, 2011. – 464 с.
Голубев А.В., Багдатьева Е.Е. Долгосрочное прогнозирование движения численности фитофагов// Лесной вестник, 2013; № 6, с 22-24.
Исаев А.С., Хлебопрос Р.Г., Кондаков Ю.П., Недорезов Л.В., Киселев В.В., Суховольский В.Г. Популяционная динамика лесных насекомых М.: Наука, 2001.- 374 с.
Ким Д.П. Теория автоматического управления. Т. 1. Линейные системы. М.: ФИЗМАТЛИТ, 2007. – 312 с. Пальникова Е.Н., Свидерская И.В., Суховольский В.Г. Сосновая пяденица в лесах Сибири. Экология, динамика
численности, влияние на насаждения. Новосибирск: Наука, 2002. - 254 с.
Пальникова Е.Н., Суховольский В.Г., Тарасова О.В. Пространственно-временная когерентность популяционной динамики лесных насекомых-филлофагов// Евразийский энтомологический журнал. – 2014. - т.13. - № 3. - С. 228-236.
Поляк Б.Т., Щербаков П.С. Робастная устойчивость и управление. М.: Наука, 2002. – 303 с.
Baltensweiler W. Zeiraphera griceana Hubner (Lepidoptera, Tortricedae) in the European Alps. A contribution to the problem of cycles// Can. Entomol.. – 1964. - v. 96. - N 5. - P. 792-800.
Baltensweiler W., Fischlin A. The larch budmoth in the Alps// Dynamics of Forest Insect Populations: Patterns, Causes, Implication. N. Y.: Plenum Press, 1988. - P. 331-351.
Schwerdtfeger F. Untersuchungen uber der eisen Bestand fon Kiefernspanner (Bupalus piniarius L.), Forleule (Panolis flammea Schiff.) und Kiefernswarmer (Hyloicus pinastri L.)// Zeitschrift fur angew. Entomol. - 1952. – 34. - № 2. – S. 216-283.
177