


Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ДИНАМИКА ГОРИЗОНТАЛЬНОЙ СТРУКТУРЫ ОСИНОВОГО ДРЕВОСТОЯ НА ПОСТОЯННОЙ ПРОБНОЙ ПЛОЩАДИ В ЧЕРНЕВОМ ПОЯСЕ ЗАПАДНОГО САЯНА
Овчинникова Н.Ф.1, Овчинников А.Е.2
1ФГБУН «Институт леса им. В.Н. Сукачева СО РАН», Красноярск, Россия
2ФГАОУВПО «Сибирский федеральный университет», Красноярск, Россия
nf@ksc.krasn.ru
Аннотация: С помощью статистического анализа выявлены и показаны особенности динамики горизонтальной структуры производного осинового древостоя по склону на постоянной пробной площади в черневом поясе Западного Саяна.
Структура древостоя в основном определяет всю ценотическую структуру лесного фитоценоза и влияет на интенсивность биогеоценотических процессов, эффективность продуцирования и депонирования органического вещества, устойчивость и биосферные функции древесных ценозов.
Динамику структуры осинового древостоя изучали на постоянной пробной площади (ППП), размером 30х40м. Все деревья, подрост и подлесок на ППП были закартированы, а особи высотой более 1,3 м и пронумерованы, что исключало ошибку их определения при периодических сплошных перечетах. У деревьев высотой 2,5 м измеряли высоту и диаметр ствола, размеры кроны, отмечали особенности онтогенеза. В Базе данных 2968 записей (Овчинникова, Овчинников, 2011). Средние древостоя – высоту и диаметр, площадь поперечного сечения и объем ствола. рассчитывали для каждого учета по общепринятым в лесной таксации методикам (Анучин, 1982).
Для описания горизонтальной структуры ППП была условно разбита на полосы: 10 шт. вдоль склона (с севера на юг) - шириной 4 м и 10 шт. поперек склона (с запада на восток) - шириной 3 м. В результате было выделено 100 прямоугольных элементарных участков с координатами по осям Х0-Х30 и У0-У40 и площадью 12 м2 каждый. Определение количества деревьев и подроста на каждом участке, и представление данных в виде матрицы дало возможность получить их частные распределения по годам учетов.
Проведенный перед закладыванием ППП на вырубке анализ размещения пней, сохранившегося тонкомера и подроста предварительных генераций показал, что горизонтальная структура срубленного коренного древостоя характеризовалась относительно равномерным размещением по площади деревьев и подроста, отражая тем самым однородность экологических условий места изучения (Ермоленко, 1981). Нарушения напочвенного и почвенного покрова при рубке темнохвойного древостоя стимулировали появление корнеотпрысковой поросли у единичных деревьев осины. Через 26 лет после рубки на ППП сформировался высокополнотный древостой I бонитета с составом по густоте 53Ос36П9Б2К и по запасу – 93Ос12П4Б1К.
Из-за выраженной эдификаторной роли верхнего яруса древостоя в таблице приведены результаты статистической обработки матриц густоты деревьев и запасов древесины на выделенных элементарных участках ППП только этого яруса. То есть, представлена динамика естественного изреживания древесного яруса и возрастание объема накопленной стволовой древесины с увеличением возраста насаждения.
Из таблицы видно, что средняя густота и запас на полосе (М) меняется со временем, т.е. с возрастом древостоя. Стандартные отклонения (δ), указывающие на разбросанность значений рядов распределения численности деревьев и их запасов в полосах, расположенных вдоль или поперек склона, имеют существенные различия между собой.
При этом значения δ для рядов распределения количества деревьев верхнего яруса в полосах, расположенных вдоль склона, в первые два учета почти в 2 раза меньше, чем в
130

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
полосах, расположенных поперек склона, а в последующие учеты – почти в 1,5. Поскольку различия значений δ имеются во все годы наблюдений, то было целесообразно определить коэффициент корреляции (r), характеризующий линейную связь между случайными величинами.
Таблица – Динамика густоты и запаса верхнего яруса древостоя и их статистических показателей в зависимости от положения на склоне
|
|
Густота |
|
|
|
Запас |
|
||
Год |
М, шт. на |
δ и t * |
|
|
М, м3 на |
δ и t |
|
|
|
полосе |
по склону |
|
|
полосе |
по склону |
|
|||
|
|
вдоль |
поперек |
|
вдоль |
поперек |
|||
1976 |
32,0 |
4,99 |
(20,3) |
9,73 |
(10,4) |
2,46 |
0,55 |
(14,06) |
0,84 (9,29) |
1980 |
27,3 |
2,83 |
(30,5) |
7,82 |
(11,04) |
3,09 |
0,72 |
(11,04) |
1,07 (9,12) |
1985 |
18,6 |
3,53 |
(16,66) |
5,13 |
(11,47) |
3,76 |
0,97 |
(11,47) |
1,68 (7,07) |
1990 |
15,6 |
3,37 |
(14,64) |
4,58 |
(10,77) |
4,1 |
1,16 |
(10,77) |
1,45 (8,93) |
1997 |
12,4 |
3,1 (12,65) |
4,53 |
(8,66) |
5,05 |
1,46 |
(8,66) |
3,87 (4,12) |
|
2008 |
7,9 |
2,28 |
(10,96) |
3,2 (7,62) |
5,38 |
2,03 |
(7,62) |
2,19 (7,77) |
|
*Критерий Стьюдента, табличное значение t при уровне значимости 0,01 и числе степеней свободы 9 составляет 2,82.
Определение r между густотой верхнего яруса древостоя в 1976 г. и его густотой в последующие годы показало, что в элементарных полосах, расположенных вдоль склона, эти коэффициенты составляют: в 1976 г. – 1,0, в 1986 г. – 0,47, в 1997 г. – 0,27 и 2008 г. – 0,67, а в элементарных полосах, расположенных поперек склона, соответственно, 1,0, 0,93, 0,87 и
0,65.
Связь между первоначальной густотой верхнего яруса древостоя и величиной отпада
впоследующие годы имела иной характер: коэффициент корреляции в элементарных полосах, расположенных вдоль склона – в 1976 г. 1,0, в 1986 г. 0,79, в 1997 г. 0,83 и 2008 г. 0,89, а в элементарных полосах, расположенных поперек склона – в 1976 г. 1,0, в 1986 г. 0,96,
в1997 г. 0,93 и в 2008 г. 0,94.
Поскольку при α > 0,05 и числе коррелируемых пар 10 критическое значение r составляет 0,632, то можно утверждать, что степень снижения густоты верхнего яруса древостоя с возрастом почти функционально определяется густотой яруса на начало исследований, при этом неравномерность горизонтальной структуры яруса постепенно снижается. Это согласуется с литературными данными о ходе изреживания древостоев с разной начальной густотой (Кузьмичев, 2013).
Использованный для анализа данных длительного мониторинга природного объекта подход позволил выявить и показать особенности динамики распределения и роста древесной растительности по склону. Влияние рельефа на структуру древостоя проявилось на сравнительно небольшой площади – 30 на 40 м, при крутизне менее 5о. Уменьшение показателей численности и роста деревьев и подроста вверх по склону необходимо учитывать при исследованиях лесных экосистем и их моделировании.
Литература
Анучин Н.П. Лесная таксация. – М.: Лесн. пром-сть. 1982. – 552 с.
Ермоленко П.М. Формирование состава хвойно-лиственных молодняков на вырубках кедровников в черневом поясе Западного Саяна // Формирование и продуктивность древостоев. – Новосибирск: Наука, Сиб. отдние, 1981. – С. 53-70.
Кузьмичев В.В. Закономерности динамики древостоев. – Новосибирск: Наука. 2013. – 208 с.
Овчинникова Н.Ф., Овчинников А.Е. Учеты деревьев на постоянных пробных площадях Красноярского края. Свидетельство о государственной регистрации базы данных №2011620706. зарег. 30.09.2011.
131
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ОЦЕНКА ПЕРВИЧНОЙ ВАЛОВОЙ И НЕТТО ПРОДУКЦИИ ЛЕСОВ ПО ВЕЛИЧИНЕ ПОГЛОЩЕННОЙ ФОТОСИНТЕТИЧЕСКИ АКТИВНОЙ РАДИАЦИИ (ФАР)
Ольчев А.В.
1 Институт Проблем Экологии и Эволюции имени А. Н. Северцова Российской академии наук, Москва, Россия
aoltche@yandex.ru
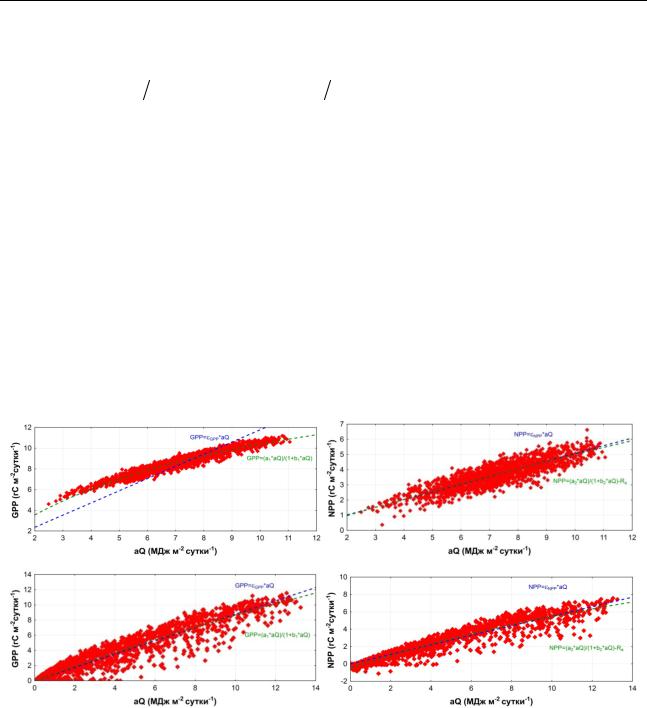
Аннотация: Для влажных тропических лесов Индонезии и еловых лесов Центральной Европы выявлены различия в зависимости интегральных месячных и суточных значений первичной валовой продукции (GPP) от величины поглощенной растительностью фотосинтетически активной радиации, ФАР (aQ). Если зависимость GPP от aQ для влажных тропических лесов имеет четко выраженный нелинейный характер, то для еловых лесов она близка к линейной. Эффективность использования растениями ФАР на GPP, для влажного тропического леса на 36% превосходит значения этого параметра для елового леса.
1. Введение
Методика расчетов среднесуточных и среднемесячных значений первичной валовой (GPP) и нетто (NPP) продукции по величине поглощенной растительным покровом ФАР (aQ) основана на гипотезе о наличии линейной зависимости между GPP и NPP растительного покрова и aQ. В качестве коэффициента пропорциональности при этом используется величина, характеризующая эффективность использования растениями ФАР
для GPP и NPP (εGPP, εNPP). Эти величины для каждого отдельного вида растительности рассматриваются как функции температуры, влажности почвы и фенологической фазы развития растения. В настоящее время данный подход нашел широкое применение для оценки GPP и NPP растительных сообществ в региональном и глобальном масштабе.
Величины εGPP и εNPP обычно оцениваются экспериментально с использованием данных прямых измерений потоков СО2 над растительным покровом (метод ковариации турбулентных пульсаций). Расчет GPP и NPP проводится с помощью, как различных методических подходов, так и с применением математических моделей.
Врамках проведенного исследования была выполнена проверка классического подхода для определения GPP и NPP по величине aQ на примере влажных тропических лесов в Индонезии и еловых лесах Центральной Европы.
2.Методы
Вкачестве основных природных объектов для изучения зависимости GPP и NPP от aQ в исследовании были выбраны вечнозеленые влажные тропические леса национального парка Лоре Линду, расположенного в центральных районах о. Сулавеси в Индонезии, а также еловые спелые леса, произрастающие в центральной Германии в Золлинговском
национальном парке. Измерения потоков СО2 проводились с помощью метода ковариации турбулентных пульсаций (eddy covariance). Расчеты по математической модели Mixfor-SVAT
использовались для восстановления непрерывных рядов данных по потокам СО2 в исследуемых лесных экосистемах в периоды, в которые измерения не проводились или определялись с погрешностями в силу низкой турбулентности приземного слоя, а также
расчетов GPP, NPP и aQ. Для анализа использовались значения GPP, NPP и aQ, осредненные за сутки и за месяц. Для параметризации зависимости GPP и NPP от aQ использовался как классический подход, основанный на линейной зависимости GPP и NPP от aQ
GPP=εGPP aQ NPP=εNPP aQ , |
(1) |
132
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
где εGPP, εNPP определяли эффективность использования растениями ФАР для GPP и NPP, так и подход, предложенный Монси и Саеки (Monsi, Saeki 1953) и позволяющий описать нелинейный характер зависимости GPP и NPP от aQ:
GPP=(a1 aQ) (1+b1 aQ) NPP=(a2 aQ) (1+b2 aQ)-Ra . |
(2) |
Параметры a1 и a2 в (2) определяют углы наклона касательных к графикам функций в точках их пересечения с осями ординат (aQ=0) и эквиваленты величинам εGPP и εNPP в (1), а
b1 и b2 характеризуют степень искривленности кривых (2). При b1, b2 → 0 нелинейные уравнения (2) превращаются в линейные (1). Ra - скорость автотрофного дыхания растений.
3. Основные результаты
Анализ зависимости интегральных месячных и суточных сумм GPP и NPP от aQ для влажного тропического леса показывает, с одной стороны, хорошо прослеживающийся нелинейный характер зависимости GPP и от aQ, а с другой стороны, близкую к линейной зависимость NPP от aQ (Рисунок 1) (Ibrom et al. 2008). Для елового леса отмечается близкая к линейной зависимость GPP и NPP от aQ как для среднемесячных, так и для
среднесуточных значений. Значение εGPP для влажного тропического леса (εGPP=1.20 гС МДж-1) на 36% превосходит εGPP для елового леса в умеренных широтах (εGPP=0.88 гС МДж- 1). Для εNPP прослеживается обратная тенденция - в случае елового леса значение εNPP выше,
чем для влажного тропического леса (εNPP=0.55 и 0.51 гС МДж-1 - для елового и влажного тропического леса, соответственно).
Рисунок 1. Зависимость дневных сумм GPP и NPP от aQ для влажного тропического леса в Барири в Индонезии (вверху) и для елового спелого леса в Центральной Германии
(внизу).
Исследование выполнено при поддержке гранта Российского Научного Фонда (РНФ
14-27-00065).
Литература
Ibrom A., Oltchev A., June T., Kreilein H., Rakkibu G., Ross Th., Panferov O., Gravenhorst G. Variation in photosynthetic light-use efficiency in a mountainous tropical rain forest in Indonesia // Tree Physiology. - 2008. - V. 28. - P. 499–508
Monsi M., Saeki T. Ueber den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung fur die Stoffproduktion // Japanese Journal of Botany. - 1953. - V. 14. - P. 22–52
133