

УЧЕБНАЯ ЛИТЕРАТУРА
.ДЛЯ МЕДИЦИНСКИХ ВУЗОВ
ВСЕ ФАКУЛЬТЕТЫ
Н.А. АГАДЖАНЯН, Л.З. ТЕЛЬ, В.И. ЦИРКИН, С.А. ЧЕСНОКОВА
ФИЗИОЛОГИЯ ЧЕЛОВЕКА
УЧЕБНИК
ИЗДАНИЕ ЧЕТВЕРТОЕ
Под редакцией академика РАМН Н.А. Агаджаняна, профессора В. И. Циркина
Рекомендовано Проблемной учебно-методической комиссией по нормальной физиологии при Всероссийском учебно-научно-методическом центре по непрерывному медицинскому и фармацевтическому образованию Минздрава РФ,
Проблемной комиссией РАМН «Эколого-физиологические проблемы адаптации»
и головным советом «Здравоохранение и экология человека» Министерства общего
и профессионального образования в качестве учебника для студентов вузов,
специализирующихся в области медицины, биологии и валеологии
![]()
Москва - МЕДИЦИНСКАЯ КНИГА Н. Новгород * Издательство НГМА
2003
ББК 28. 903 Ц23
Агаджанян Н.Л., Тель Л.З., Циркин В.И., Чеснокова С.А. ФИЗИОЛОГИЯ
ЧЕЛОВЕКА. — М.: Медицинская книга, Н. Новгород: Издательство НГМА,
2003.-528 с: илл.
Учебник в доступной форме отражает современное состояние физиологии человека — фундаментальной дисциплины в образовании будущего врача, биолога, валеолога. В книге представлены все основные разделы физиологии, предусмотренные соответствующими государственными образовательными стандартами. Многолетний опыт преподавания курса нормальной физиологии студентам медицинского факультета Российского Университета дружбы народов, Челябинского, Кировского и Акмолинского медицинских институтов позволил авторам учебника при сохранении должного научного уровня изложить ясно, четко и сравнительно просто сложнейшие вопросы физиологии человека. Об этом свидетельствует популярность предыдущих изданий учебника среди студентов и преподавателей вузов (факультетов) медицинского и биологического профиля. Учебник, кроме того, может быть полезным для учащихся медицинских колледжей и инновационных школ с углубленным изучением биологии, а также для студентов и аспирантов, специализирующихся в области экологии человека, психологии, физической культуры и спорта, ветеринарии.
Рецензенты:
Ю.В. Урываев, д.м.н., профессор, зав. кафедрой нормальной физиологии
химико-фармацевтического факультета Московской медицинской академии;
В.Н. Кобрин, д.м.н., профессор кафедры нормальной физиологии
'Российского государственного медицинского университета.
По вопросам приобретения книги
обращайтесь по телефонам:
(095) 189-99-35 (8313)25-57-11
ВОЗМОЖНО ПОЛУЧЕНИЕ КНИГИ СО СКЛАДА В МОСКВЕ И ДОСТАВКА В РЕГИОНЫ
ISBN 5-86093-061-5
Коллектив авторов, 2003
СПИСОК ИСПОЛЬЗОВАННЫХ СОКРАЩЕНИЙ
АР — адренорецспторы
АРФ — абсолютная рефрактерная фаза
АХ — ацетилхонин
АТФ — аденозинтрифосфат
АХЭ —ацетилхолинэстераза
АКТГ — адренокортикотропный гормон
АД — артериальное давление
АДФ — аденозиндифосфат
АДГ — антидиуретическнй гормон
БКГ — баллнстокардиография
БО — буферные основания
ВПСП— возбуждающий постсинаптический
потенциал
BE — буфер-эксцесс ВНС — вегетативная нервная система ВО — буферные основания ВИП — вазоактивный интестиадьный
пептид
BOO — величина основного обмена ГАМК— гамма-аминомасляная кислота ГИП — гастро-ингибнрующий пептид ГМК — гладкомышечные клетки ДО — дыхательный объем ДЕ — дыхательная единица ДК — дыхательный коэффициент ЖКТ — желудочно-кишечный тракт КУД — критический уровень
деполяризации
КОМТ— катехоламин-О-метилтрансфераза КЩР — кислотно-щелочное равновесие КЭК — калорический эквивалент
кислорода КСФ — колониестимулирующий фактор
ЛГ — лютеинизирующий гормон МП — мембранный потенциал МАО — моноаминоксидаза МСГ — меланоцитстимулирующий
гормон
МОД — минутный объем-дыхания МОК — минутный объем крови МВЛ — максимальная вентиляция легких НА — норадреналин ОЕЛ — общая емкость легких ООЛ — общий объем легких ОРФ — относительная рефрактерная фаза ПКП — потенциал концевой
пластинки ПРЛ — пролактин ПД — потенциал действия СКК — стволовая кроветворная клетка СТГ — соматотропный гормон СО — систолический объем СР — саркоплазматическийретикулюм ТПСП-—тормозной постсинаптический
потенциал
ТТГ — тиреотропный гормон ФСГ — фоллнкулостимулирующий
гормон
ФС — функциональная система ФОЕ — функциональная остаточная
емкость
ХР — холинорецептор цАМф— циклический АМФ ЭКГ — электрокардиограмма ЭЭГ — электроэнцефалограмма ЮГА — юкстагломерулярный аппарат
Глава 1
ТРАНСПОРТ ВЕЩЕСТВ. ЭЛЕКТРОГЕНЕЗ. ВОЗБУДИМЫЕ ТКАНИ
СОВРЕМЕННОЕ ПРЕДСТАВЛЕНИЕ О СТРОЕНИИ И ФУНКЦИИ МЕМБРАН
Детальное знакомство со строением и функциями биологических мембран дается в курсах гистологии, биофизики. С позиций физиологии важны следующие детали: всякая плазматическая мембрана имеет два слоя фосфолипидов. В них встроены белки (интегральные), а на их поверхности имеются периферические белки и углеводы. Последних много, особенно в чнтероцитах, они образуют гликокаликс, выполняющий определенную функцию. В частности у энтероцитов он используется как фильтр при всасывании веществ. Белки играют важную роль: это ферменты (например, участвующие в гидролизе АТФ), переносчики, ионные каналы, рецепторы. Все белки мембраны синтезируются в эндоплазматичес-ком ретикулюме, а затем направляются в аппарат Гольджи, откуда они распределяются на соответствующие участки мембраны.
Мембрана играет большую роль в жизнедеятельности клетки. Одной из функций мембраны является барьерно-транспортная: мембрана определяет потоки веществ, идущих через нее, микросреду, т. е. состав цитоплазмы. Мембрана принимает участие в генерации ПД (потенциала действия), в генерации энергии (например, мембраны митохондрий). Мембраны осуществляют функцию межклеточного взаимодействия — например, передачу сиг-пала от одного нейрона к другому.
ТРАНСПОРТ ВЕЩЕСТВА ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Классификация. Различают прямой и опосредованный транспорт. Прямой — без участия переносчиков, опосредованный — с их участием. Например, перенос глюкозы с участием переносчика.
Опосредованный транспорт осуществляется с затратой энергии (активный транспорт) или без затраты энергии (облегченная диффузия). Прямой транспорт всегда идет по типу пассивного транспорта.
Из сказанного ясшо, что второй вариант классификации — это выделение двух основных видов транспорта веществ — пассивного и активного.
Третий вариант классификации — транспорт с изменением архитектуры мембраны (эк-зоцитоз, эндоцитоз) или без изменения структуры мембраны (все остальные виды транспорта).
Четвертый вариант классификации — это транспорт, сопряженный с переносом двух веществ (котранспорт), который может протекать по типу симлорта (два вещества идут в одном направлении — например Na + глюкоза) или по типу антипорта (одно вещество идет в клетку, второе — из клетки или наоборот — Na и К).
Антипод котранспорта — обычный транспорт, или унипорт, т. е. когда переносится одно вещество, например, молекулы глюкозы.
Пассивный транспорт. Различают два его вида — простую диффузию и облегченную диффузию. Простая диффузия идет в соответствии с законом Фика — по химическому или электрохимическому градиенту. Например, в клетке натрия 14 ммоль, а в среде — 140 ммоль, в этом случае пассивный поток должен быть направлен в клетку. В живых системах
этот вид транспорта используется ограниченно. Кислород, углекислый газ, вода—они могут таким образом осуществлять переход из клетки в среду или наоборот. Важно, что для пассивной простой диффузии вещество должно быть жирорастворимым. Гидрофильные вещества пройти в фосфолипидном слое мембраны не могут. В целом, затраты энергии на этот процесс не происходит, так как диффузия идет по градиенту.
Облегченная диффузия проходит по двум вариантам — с участием переносчиков или при наличии специализированных каналов. Обнаружено, что в мембранах, имеются специальные белки-переносчики, которые, специфически связываясь с переносимой молекулой, способствуют ее переносу по градиенту концентрации. Таким образом, затраты энергии не происходит, а путь через мембрану облегчается за счет наличия специфического переносчика. Специфичность проявляется, например, в том, что D-изомеры Сахаров транспортируются таким образом, а L-изомеры нет, или L-аминокислоты транспортируются, а D-амино-кислоты нет.
Рис. 1. Схема строения мембраны возбудимой ткани.
— слой билипидоа и каналы для ионов Na;
— канал для ионов Na а закрытом состоянии;
— «ворота» открыты;
— «ворота» инактивации закрыты;
— селективный фильтр.
И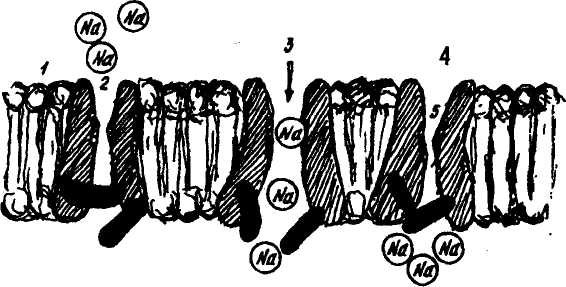 онные
каналы — это интегральные
белки мембраны, которые выполняют
функцию транспортирующей
частицы для соот-ветствующего
иона. Селективность
канала обеспечивается
за счет геометрии
канала (диаметр устьев,
диаметр селективного
фильтра), за счет внут-риканально
расположенных
заряженных частиц (например,
для катион-пропускающих
каналов — анионные
частицы). Каждый ионный канал имеет
устье,
селективный фильтр,
ворота и механизм
управления воротами.
Часть каналов управляется за счет
разности потенциалов на мембране
(потенциал-зависимые
ионные каналы) — для этого рядом с
каналом имеется электрический сенсор,
который
в зависимости от величины мембранного
потенциала либо открывает ворота
каналов,
либо держит их закрытыми. Второй вариант
ионных каналов — рецепторуправляемые
каналы:
в этом случае ворота каналов управляются
за счет рецептора, расположенного на
поверхности мембраны: при взаимодействии
медиатора (лиганда) с этим рецептором
может происходить открытие ионных
каналов. В некоторых рецепторуправляемых
каналах между
рецептором и воротным механизмом имеется
промежуточная стадия (посредник типа
цАМФ,
протеинкиназы и т. п.)-
онные
каналы — это интегральные
белки мембраны, которые выполняют
функцию транспортирующей
частицы для соот-ветствующего
иона. Селективность
канала обеспечивается
за счет геометрии
канала (диаметр устьев,
диаметр селективного
фильтра), за счет внут-риканально
расположенных
заряженных частиц (например,
для катион-пропускающих
каналов — анионные
частицы). Каждый ионный канал имеет
устье,
селективный фильтр,
ворота и механизм
управления воротами.
Часть каналов управляется за счет
разности потенциалов на мембране
(потенциал-зависимые
ионные каналы) — для этого рядом с
каналом имеется электрический сенсор,
который
в зависимости от величины мембранного
потенциала либо открывает ворота
каналов,
либо держит их закрытыми. Второй вариант
ионных каналов — рецепторуправляемые
каналы:
в этом случае ворота каналов управляются
за счет рецептора, расположенного на
поверхности мембраны: при взаимодействии
медиатора (лиганда) с этим рецептором
может происходить открытие ионных
каналов. В некоторых рецепторуправляемых
каналах между
рецептором и воротным механизмом имеется
промежуточная стадия (посредник типа
цАМФ,
протеинкиназы и т. п.)-
Селективность ионных каналов во многом определяется геометрией канала. Например, кристаллический диаметр иона натрия — 0,19 нм, вместе с гидратной оболочкой его диаметр становится около 0,3 нм. Устье канала 0,3—0,5 нм. Для того, чтобы пройти через канал (главное — через селективный фильтр), ион натрия или другой ион должен освободиться от гидратной оболочки и только в «голом» виде может пройти через канал. Слишком большой ион не может войти в канал, слишком маленький не способен отдать гидрат-ную оболочку в селективном фильтре, поэтому не может выскочить из канала.
Натриевые каналы имеют устья, селективный фильтр, воротный механизм. Ворота у них двух типов — актнвацнонные (м-ворота) и ннактивационные (h-ворота). В условиях покоя (МП=80 мВ, например), активационные ворота закрыты, но готовы в любую минуту
открыться, а ннактивационные ворота открыты. При снижении МП (например, до 60 мВ) активационные ворота открываются и впускают ионы натрия в клетку, но вскоре начинают закрываться инактивационные ворота (происходит инактивация натриевых каналов). Некоторое время спустя закрываются активационные ворота, открываются инактивационные ворота, и канал готов к новому циклу. Канал блокируется тетродотоксином, местными анестетиками (новокаином, другими веществами). Это используется в медицинской практике.
Калиевые каналы тоже достаточно селективны — в основном пропускают ионы калия. Они блокируются тетраэтиламмонием. Процессы инактивации у них выражены слабо.
Кальциевые каналы — имеют все атрибуты ионного канала (устья, воротный механизм, фильтр). Блокируются ионами марганца, никеля, кадмия (двухвалентные ионы), а также лекарственными веществами — верапамилом, нифедипином, дильтиаземом, которые используются в клинической практике.
Активный транспорт: различают первично-активный транспорт, при котором энергия затрачивается на перенос данного вещества против градиента его концентрации, а также вторично-активный транспорт, при котором энергия на перенос данного вещества (например, молекулы глюкозы) используется за счет механизма переноса другого вещества (например, натрия).
Первично-активный транспорт широко представлен в организме. Это калий-натриевый насос, натрий-водородный обменный механизм, натрий-кальциевый обменный механизм, кальциевый насос и т. д. Суть его состоит в том, что в мембране имеется переносчик, обладающий АТФ-азной активностью, т. е. он способен расщеплять АТФ и высвобождать энергию, которая и затрачивается на перенос вещества. Конкретно: калий-натриевый насос «выкачивает» из клетки ионы натрия, а «вкачивает» ионы калия (против градиента концентрации). Чтобы осуществить перенос натрия из клетки в среду, переносчик (калий-натриевая АТФ-аза) внутри клетки соединяется с ионами натрия, в результате активируется АТФ-аз-ная активность переносчика, происходит гидролиз АТФ, это вызывает высвобождение энергии, в результате *— переносчик каким-то образом (типа качели?) переносит натрий в среду. Здесь он теряет сродство к натрию» но приобретает сродство к кадию а присоединяет его ионы. В результате — меняется конформация переносчика, и он (каким-то образом?) вновь возвращается к внутренней поверхности мембраны, внося в клетку ионы калия. Здесь вновь он теряет сродство к ионам калия, но приобретает сродство к ионам, натрия, и цикл повторяется. Насос ингибируется уабаином (строфантин G). Конкретные механизмы работы насоса еще во многом не ясны.
Вторично-активный транспорт. В основном представлен в энтероцитах, в эпителии почек. Суть его состоит в следующем (на примере переноса молекулы глюкозы). Молекула глюкозы должна войти в клетку, где ее концентрация намного выше, чем в среде. Для того, чтобы это произошло, необходимы затраты энергии. Но тратится энергия, которая ранее была затрачена на перенос натрия. Дело в том, что в этой клетке создаются за счет работы калий-натриевого насоса низкие концентрации натрия. При наличии высоких концентраций натрия в среде — натрий будет стремиться войти в клетку (по градиенту). Итак, молекула глюкозы присоединяется к специфическому переносчику, к которому подсоединяется ион натрия. В результате градиента концентрации (для натрия) этот «комбайн» (переносчик + глюкоза + ион натрия) переносится внутрь клетки, где глюкоза и натрий отщепляются от переносчика, а переносчик «уходит» вновь совершать свою работу. Натрий откачивается помпой, а глюкоза покидает клетку с другой стороны уже по градиенту концентрации (облегченная диффузия по типу «перенос с участием переносчика»).
В целом, представленные примеры демонстрируют наличие в живых системах котранс-порта, когда одним механизмом транспортируется одновременно две молекулы. В случае калий-натриевого насоса имеет место антипорт (калий идет в одну сторону, натрий в противоположную), в случае вторично-активного транспорта имеет место симпорт.
Эндоцитоз и экэоцитоз. Это варианты транспорта, при которых меняется архитектура мембран. Он осуществляется с затратой энергии. Эндоцитоз — это введение крупномолекуляр-
ных частиц из среды в клетку. Один его вариант — фагоцитоз, другой ~ пиноцитоз. Пиноци-тоз представляет собой способ усвоения клеткой белковой молекулы без се предварительного гидролиза. Такой механизм, например, имеет место у новорожденных, которые с молоком матери получают антитела (иммуноглобулины), через эитероциты попадающие в организм ребенка, будучи совершенно ненарушенными и способными к выполнению своих функций.
Экзоцитоз — это выделение крупных молекул из клетки. Пример тому — выделение квантов медиатора из везикулы в синапсе.
ВОЗБУДИМЫЕ ТКАНИ И ИХ ОБЩИЕ СВОЙСТВА
Возбудимые ткани — это нервная, мышечная и железистая структуры, которые способны спонтанно или в ответ на действие раздражителя возбуждаться. Возбуждение — это генерация потенциала действия (ПД) + распространение ПД + специфический ответ ткали на этот потенциал, например, сокращение» выделение секрета, выделение кванта медиатора.
Свойства возбудимых тканей н показатели, их характеризующие:
Свойства
1. Возбудимость — способность
возбуждаться
2. Проводимость — способность проводить возбуждение, т. е. проводить ДЦ
Сократимость — способность развивать силу или напряжение при возбуждении
Лабильность — или функциональная подвижность — способность
к ритмической активности
5. Способность выделять секрет (секреторная активность), медиатор
Детальнее — см. ниже.
Показатели
Порог раздражения, реобаза, хронаксия, длительность абсолютной рефрактерной фазы, скорость аккомодации.
Скорость проведения ПД, например, у нерва она может достигать 120 м/с (около 600 км/час).
Максимальная величина силы (напряжения), развиваемая при возбуждении.
Максимальное число возбуждений в единицу времени, например, нерв способен в 1с генерировать 1000 ПД
Величина квантового выхода, обьем секрета
ЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ВОЗБУДИМЫХ ТКАНЯХ
Классификация:
Биопотенциалы — общее название всех видов электрических процессов в живых системах.
Потенциал повреждения — исторически первое понятие об электрической активности живого (демаркационный потенциал). Это разность потенциалов между неповрежденной и поврежденной поверхностями живых возбудимых тканей (мышцы, нервы). Разгадка его природы привела к созданию мембранной теории биопотенциалов.
Мембранный потенциал (МП) — это разность потенциалов между наружной и внутренней поверхностями клетки (мышечного волокна) в покое. Обычно МП, или потенциал покоя, составляет 50—80 мВ, со знаком «—» внутри клетки. При возбуждении клетки регис-грируется потенциал действия (его фазы: пик, следовая негативность, следовая позитивность) — быстрое изменение мембранного потенциала во время возбуждения.
Внеклеточно-регнстрируемый потенциал действия, внутриклеточно-регистрируемый потенциал действия — это варианты потенциалов действия, форма которых зависит от способа отведения (см. ниже).
8
Рецепторный (генераторный) потенциал — изменение МП рецепторных клеток во время их возбуждения.
Постсннаптические потенциалы (варианты: возбуждающий постсинаптический потенциал — ВПСП, тормозной постсинаптический потенциал — ТПСП, частный случай возбуждающего постсинаптического потенциала — ПКП — потенциал концевой пластинки).
Вызванный потенциал — это потенциал действия нейрона, возникающий в ответ на возбуждение рецептора, несущего информацию к этому нейрону.
ЭКГ (грамма), ЭЭГ, ЭМГ (миограмма) — соответственно — суммарная электрическая активность сердца, мозга, скелетных мышц при их возбуждении.
История — это Гальвани, Маттеучи, Дюбуа-Реймон, Бернштейн, Ходжкин, Хаксли, Катц.
Более детально все виды биоэлектрической активности будут описаны в последующем.
ЭКСПЕРИМЕНТАЛЬНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ БИОЭЛЕКТРИЧЕСКИХ ЯВЛЕНИИ
Рис. 2. Скорость распространения возбуждения по нервным волокнам разного типа.
i — схема опыта раздражения нервного ствола стимулятором (Ст) и отведение биотока из ближней точки (а) и отдаленной (б) с помощью установок, включающих электрод, усилитель, осциллоскоп (соответственно Ус и Ос), М — мышца. II — нерв, состоящий из волокон типов А, В, С. Человечки — импульсы, бегущие по волокнам с разной скоростью. Диссоциация скоростей особенно заметна на экране осциллоскопа. На графике представлены соотношения потенциалов действия волокон А (а Р,у), В, С.
Л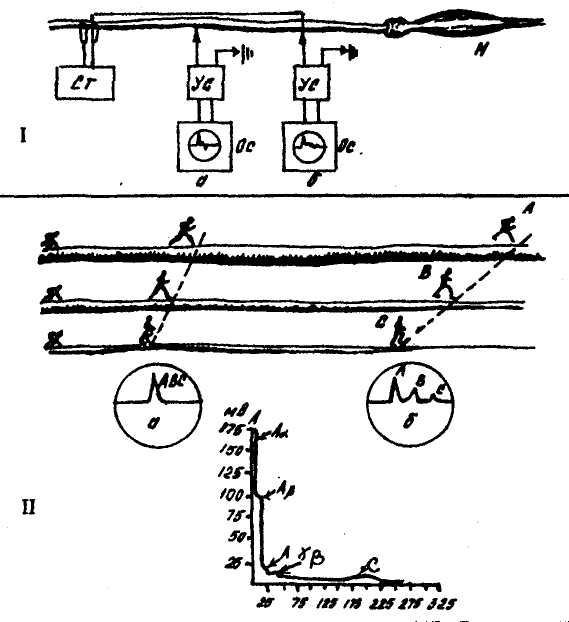 .
Гальвани был первым,
кто убедился в существовании
«живого электричества».
Его первый
(балконный) опыт состоял
в том, что препарат
задних лапок лягушек
на медном крючке был
подвешен к железному
балкону. От ветра он задевал
балконные перила,
и это вызывало сокращение
мышц. По Гальва-ви, это было результатом
замыкания цепи
тока, в результате
чего «живое электричество»
вызывало
сокращение. Вольта (итальянский
физик) опроверг
такое объяснение. Он
полагал, что сокращение обусловлено
наличием «гальванической пары»
— железо-медь. В ответ Гальвани поставил
второй опыт
(опыт без металла),
который доказывал
идею автора: набрасывался
нерв между поврежденной
и неповрежденной
поверхностями
мышцы и в ответ — сокращение
интактной мышцы.
.
Гальвани был первым,
кто убедился в существовании
«живого электричества».
Его первый
(балконный) опыт состоял
в том, что препарат
задних лапок лягушек
на медном крючке был
подвешен к железному
балкону. От ветра он задевал
балконные перила,
и это вызывало сокращение
мышц. По Гальва-ви, это было результатом
замыкания цепи
тока, в результате
чего «живое электричество»
вызывало
сокращение. Вольта (итальянский
физик) опроверг
такое объяснение. Он
полагал, что сокращение обусловлено
наличием «гальванической пары»
— железо-медь. В ответ Гальвани поставил
второй опыт
(опыт без металла),
который доказывал
идею автора: набрасывался
нерв между поврежденной
и неповрежденной
поверхностями
мышцы и в ответ — сокращение
интактной мышцы.
В настоящее время существуют два основных метода регистрации
биопотенциалов; внеклеточный и внутриклеточный. Внеклеточный способ — это отведение разности потенциалов между двумя точками ткани, органа. Варианты — монополярное отведение (один электрод заземлен), биполярное отведение (оба электрода активные). Контактный способ — электроды непосредственно соприкасаются с объектом исследования, дистантный (например, при ЭКГ-графии) — между объектом исследования и электродами имеется среда. В целом, при внеклеточном методе отводится лишь часть потенциала. Мембранный потенциал замерить нельзя.
Внутриклеточный способ: один электрод — в среде, второй (стеклянная пипетка) — вводится внутрь клетки. Регистрируется разность потенциалов между наружной и внутренней поверхностями мембраны. Пипетка предварительно заполняется раствором хлористого калия.
МЕМБРАННЫЙ ПОТЕНЦИАЛ И ЕГО ПРОИСХОЖДЕНИЕ
МП, или потенциал покоя, — это разность потенциалов между наружной и внутренней поверхностями мембраны в условиях покоя. В среднем у клеток возбудимых тканей он достигает 50—80 мВ, со знаком «—» внутри клетки. Обусловлен преимущественно ионами калия. Как известно, в клетках возбудимых тканей концентрация ионов калия достигает 150 ммоль/л, в среде — 4—5 ммоль (ионов калия намного больше в клетке, чем в среде). Поэтому по градиенту концентрации калий может выходить из клетки, и это происходит с участием калиевых каналов, часть которых открыта в условиях покоя. В результате из-за того, что мембрана непроницаема для анионов клетки (глутамат, аспартат, органические фосфаты), на внутренней поверхности клетки образуется избыток отрицательно заряженных частиц, а на наружной — избыток положительно заряженных частиц. Возникает разность потенциалов. Эта ситуация описывается уравнением Нернста:
п - ПX л* Кв ** F ** Ki ■
Чем выше концентрация калия в среде — тем меньше это отношение, тем меньше величина мембранного потенциала. Однако расчетная величина, как правило, ниже реальной, Например, по расчетам МП должен быть -90 мВ, а реально -70 мВ. Это расхождение обусловлено тем, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В частности, известно, что натрия больше в среде (140 ммоль/л против 14 ммоль/л внутриклеточной). Поэтому натрий может войти в клетку. Но большая часть натриевых каналов в условиях покоя закрыта. Поэтому в клетку входит лишь небольшая часть ионов натрия. Но и этого достаточно, чтобы хотя бы частично компенсировать избыток анионов. Ионы хлора, наоборот, входят в клетку (частично) и вносят отрицательные заряды. В итоге величина мембранного потенциала определяется в основном калием, а также натрием и хлором. Формула Гольдмана-Ходжкина (см. в учебнике) описывает это явление.
Для того чтобы МП поддерживался на постоянном уровне, необходимо поддержание ионного гетерогенитета — ионной асимметрии. Для этого, в частности, служит калий-натриевый насос (и хлорный), который восстанавливает ионную асимметрию, особенно после акта возбуждения. Доказательством калиевой природы МП является наличие зависимости: чем выше концентрация калия в среде, тем меньше величина МП. Для дальнейшего изложения важно понятие; деполяризация (уменьшение МП, например, от минус 90 мВ до минус 70 мВ) и гйперполяризация — противоположное явление.
ПОТЕНЦИАЛ ДЕЙСТВИЯ
Потенциал действия — это кратковременное изменение разности потенциала между наружной и внутренней поверхностями мембраны (или между двумя точками ткани), возникающее в момент возбуждения. При регистрации потенциала действия с помощью микроэлектродной техники наблюдается типичный пикообразный потенциал. В нем выделяют
следующие фазы или компоненты:
10
Локальный ответ — начальный этап деполяризации.
Фазу деполяризации — быстрое снижение мембранного потенциала до нуля и переза рядка мембраны (реверсия, или овершут).
Фазу реполяризации — восстановление исходного уровня мембранного потенциала; в ней выделяют фазу быстрой реполяризации и фазу медленной реполяризации; в ейою оче редь, фаза медленной реполяризации представлена Следовыми процессами (потенциалами): следовая негативность (следовая деполяризация) и следовая позитивность (следовая гипер поляризация). Амплитудно-временные характеристики потенциала действия нерва, скелет ной мышцы таковы: амплитуда потенциала действия 140—150 мВ; длительность пика по тенциала действия (фаза деполяризации + фаза реполяризации) составляет 1 —2 мс, дли тельность следовых потенциалов — 10—50 мс.
Форма потенциала действия (при внутриклеточном отведении) зависит от вида возбудимой ткани: у аксона нейрона, скелетной мышцы — пикообразные потенциалы, у гладких мышц в одних случаях пикообразные, в других — платообраэные (например, потенциал действия гладких мышц матки беременной женщины — платообразный, а длительность его составляет почти 1 мшгуту). У сердечной мышцы потенциал действия имеет платообразную форму.
При внеклеточной регистрации форма потенциала иная, и она зависит от того, какой вид отведения используется — монополярный или биполярный. При биполярном отведении в покое разность потенциалов между двумя точками ткани (например, между точкой 1 и 2) отсутствует. Когда появляется возбуждение, например, распространяющееся в направлении через точку 1 к точке 2 и далее, волна возбуждения достигает вначале электрода 1 — возникает разность потенциалов, при этом область у электрода I — электронегативна по отношению к области под электродом 2. Когда волна достигает области под электродом 2, то эта область становится электронегативной по отношению к области под электродом 1. В целом возникает двухфазное изменение разности потенциалов. При монополярном отведении в условиях, когда один из электродов заземлен, потенциал действия по форме однофазный, т. к. вторая область всегда имеет постоянный потенциал, равный 0.
Амплитудные характеристики: если при внутриклеточном отведении амплитуда ПД достигает 120— 150 мВ, то при внеклеточном отведении, особенно при дистантном отведении, амплитуда ПД составляет всего 1—2 мВ (например, ЭКГ) или 100—200 мкВ (ЭЭГ). Иначе говоря, при внеклеточном отведении регистрируется только сотая часть потенциала. Важно подчеркнуть, что при внеклеточном отведении регистрируется активность множества одиночных образований, которые в определенной степени могут возбуждаться асинхронно. Это отражается на реальной форме внеклеточно отводимого потенциала. Такова, например, ситуация в отношении ЭКГ.
ПРИРОДА ПОТЕНЦИАЛА ДЕЙСТВИЯ
При исследовании ПД аксонов и сомы нервной клетки, ПД скелетной мышцы было установлено, что фаза деполяризации обусловлена значительным повышением проницаемости для ионов натрия, которые входят в клетку в начале процесса возбуждения и таким образом уменьшают существующую разность потенциала (деполяризация). Процесс этот имеет регенеративную природу — чем выше степень деполяризации, тем выше становится проницаемость натриевых каналов, тем больше входит ионов натрия в клетку и тем выше степень деполяризации. В этот период происходит не только снижение разности потенциалов до нуля, но и изменение поляризованное™ мембраны — на высоте пика ПД внутренняя поверхность мембраны заряжена положительно по отношению к наружной (явление реверсии, или овершута). Однако бесконечно этот процесс идти не может: в результате закрытия инактивациоиных ворот натриевые каналы закрываются, и приток натрия в клетку прекращается. Затем наступает фаза реполяризации. Она связана с увеличением выхода из клетки ионов калия. Это происходит за счет того, что в результате деполяризации большая часть калиевых каналов, которые в условиях покоя были закрыты, открываются и «+» заряды
11
уходят за пределы клетки. Вначале этот процесс идет очень быстро, потом — медленно, поэтому фаза реполяризации вначале протекает быстро (нисходящая часть пика ПД), а потом медленно (следовая негативность). Этот же процесс чежит в основе фазы следовой гиперполяризации. На фоне следовых потенциалов происходит активация калий-натриевого насоса. Если он работает в электронейтральном режиме (2 иона натрия вьшосятся из клетки в обмен на 2 вносимых в клетку иона калия), то на форме ПД этот процесс не отражается. Если же насос работает в электрогенном режиме, когда 3 иона натрия выносятся из клетки в обмен на 2 вносимых в клетку иона калия, то в результате на каждый такт работы насоса в клетку вносится на 1 катион меньше, чем выносится, поэтому в клетке постепенно возрастает избыток анионов, т. е. в таком режиме насос способствует появлению дополнительной разности потешдиааов. Это явление может лежать в основе фазы следовой гиперполяризации.
Если насос заблокировать, например, с помощью специфического блокатора — строфантина G (уабаин), то постепенно клетка перестает отвечать на раздражители, т. к. ионный гетерогенитет как основа электрической активности резко снизится,
Одним из доказательств того, что в аксонах, нейронах, скелетных мышцах ПД по природе натриевый, являются опыты с гипонатриевыми растворами: если ткань помещена в такой раствор, то величина амплитуды ПД снижается и тем на большую величину, чем меньше в среде содержание натрия. Блокаторы натриевых каналов, например, тетродотоксин, также приводят к ингибированию процесса генерации ПД, к снижению амплитудных и временных характеристик ПД. Анестетики за счет их способности блокировать натриевые каналы тоже вызывают нарушение возбудимости, что используется в клинической практике для блокады проведения по нерву.
В сердечной мышце природа ПД иная: процесс деполяризации обусловлен ионами натрия и кальция — эти ионы входят внутрь клетки в начале фазы деполяризация. Поэтому потенциал действия сердечной мышцы может частично уменьшаться блокаторами натриевой проницаемое га (тетродотоксином, новокаином), а частично — блокаторами кальциевой проницаемости (верапамилом, нифедипином). В настоящее время блокаторы кальциевой проницаемости используются с целью подавления активности клеток сердца в условиях клиники.
В гладких мышцах сосудов, желудка, кишечника, матки и других образований генерация; ПД связана с тем, что в момент возбуждения в клетку входят главным образом не ионы натрия, а ионы кальция. Поэтому блокаторы кальциевой проницаемости, типа верапамила, тормозят активность гладкомышечных клеток. Это используется в клинике, в частности, при лечении угрозы прерывания беременности, т. е. для ингибирования сократительной активности матки.
ЗАКОНЫ РАЗДРАЖЕНИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Прежде чем рассмотреть эти законы, необходимо представить, каким образом происходит возбуждение, т. е. какие условия должны возникнуть в возбудимой ткани, чтобы она реализовала свою способность возбуждаться. Основное условие — это снижение мембранного потенциала до критического уровня деполяризации (КУД). Любой агент, если он способен это сделать, одновременно вызывает и возбуждение ткани. Например, МП -70 мВ. КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т. е. на 20 мВ снизить ее исходный потенциал покоя. Как только МП достигнет уровня КУД, то в дальнейшем процесс (в силу регенеративности) будет продолжаться самостоятельно и приведет к открытию всех натриевых каналов, т. е. к генерации полноценного ПД. Если мембранный потенциал не достигнет этого уровня, то в лучшем случае возникнет так называемый местный потенциал (локальный ответ).
Все агенты, которые вызывают гиперполяризацию ткани, в момент воздействия не смогут вызвать возбуждение, т. к. в этом случае МП не достигает критического уровня деполяризации, а наоборот, уходит от него.
12
Три замечания:
В ряде возбудимых тканей величина мембранного потенциала по времени непостоян на — она периодически снижается и самостоятельно достигает КУД, в результате чего воз никает спонтанное возбуждение (автоматия). Это характерно для водителей ритма сердца, для некоторых гладких мышц, например, мышц матки.
Когда на ткань действует раздражитель (в подпороговой силе), то он может вызывать изменение КУД. Например, длительная подпороговая деполяризация приводит к тому, что КУД изменяется: допустим, в исходном состоянии он -составляет -50 мВ, а в результате длительной деполяризации он становится равным -40 или -30 мВ. В такой ситуации вызы вать возбуждение становится труднее. В целом, это явление получило название аккомода ции возбудимой ткани. Оно лежит в основе закона градиента (не путать с понятием «акко модация глаза»).
Для возбуждения ткани необходимо наличие внешнего раздражителя по отношению к этой ткани (исключение — ткани, обладающие автоматией). Такими раздражителями в ес тественных условиях могут быть нервный импульс, выделение медиатора. В целом, в физи ологии говорят о двух видах раздражителей — адекватных и неадекватных. Адекватные раздражители — это такие воздействия, которые «в малых дозах» способны вызвать воз буждение (ткань в процессе эволюции к ним приспособлена). Например, квант света для фото рецептора, нервный импульс для синапса. Неадекватный раздражитель тоже способен вызывать возбуждение, но для этого он должен быть использован в больших «дозах», в результате чего ткань может повреждаться.
Для того, чтобы раздражитель вызвал возбуждение, он должен быть: 1. достаточно сильным (закон силы), 2. достаточно длительным (закон времени), 3. достаточно быстро нарастать (закон градиента). Если эти условия не соблюдаются, то возбуждения не происходит. Рассмотрим подробнее эти законы раздражения и следствия, которые из них вытекают.
Закон силы. Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным — пороговым или выше порогового. Обычно под термином «порог» понимается минимальная сила раздражителя, которая способна вызвать возбуждение. Например, чтобы вызвать возбуждение нейрона при МП - -70 мВ и КУД = -50 мВ, пороговая сила должна быть равной -20 мВ. Этот закон рассматривает также зависимость амплитуды ответа возбудимой ткани от силы раздражителя (раздражитель по силе ниже пороговой величины, равен или выше ее). Для одиночных образований (нейрон, аксон, нервное волокно) эта зависимость носит название правила «все или ничего». Например, регистрируется ответ ткани — потенциал действия аксона. В качестве параметра ответа возьмем его амплитуду. Пусть величина раздражителя составляет 10 мВ, ответ отсутствует (раздражитель является допорого-вым), далее — раздражитель равен ,30 мВ — возникает ответ в виде ПД, его амплитуда равна 130 мВ. Увеличим силу раздражителя (до 50 мВ) —вновь генерируется ответ в виде потенциала действия, его амплитуда равна 130 мВ. Следующий раздражитель по силе — 100 мВ, амплитуда ПД — 130 мВ. Вот пример правила «все или ничего».
Если речь идет о целом образовании, например, нервный ствол, содержащий отдельные аксоны, или о скелетной мышце как совокупности отдельных мышечных волокон, то в этом случае каждое отдельное волокно тоже отвечает на раздражитель по типу «все или ничего», но если регистрируется суммарная активность объекта (например, внеклеточно отводимый ПД), то его амплитуда в определенном диапазоне находится в градуальной зависимости от силы раздражителя: чем больше сила раздражителя, тем больше ответ. Пример: пусть имеется нервный ствол, состоящий из 10 аксонов. Пороги раздражения для них таковы: 30 мВ — 1-й, 40 мВ — 2, 3, 4-й, 50 мВ — 5, 6, 7, 8-й и 60 мВ —'9 и 10-й аксоны. Следовательно, при 30 мВ активируется 1 аксон, при 40 мВ — 4 (1-й + 2,3,4-й), при 50 мВ — 8 (1-й + 2,3,4-й + 5,6,7, 8-й), а при 60 мВ — все 10 волокон. Таким образом, в пределах от 30 до 60 мВ имеет место градуальная зависимость. При дальнейшем увеличении силы раздражителя амплитуда суммарного ответа постоянна.
Одно важное следствие этого закона — введено понятие «порог раздражения» (минимальная сила раздражителя, способного вызвать возбуждение). Определяя этот показатель,
13
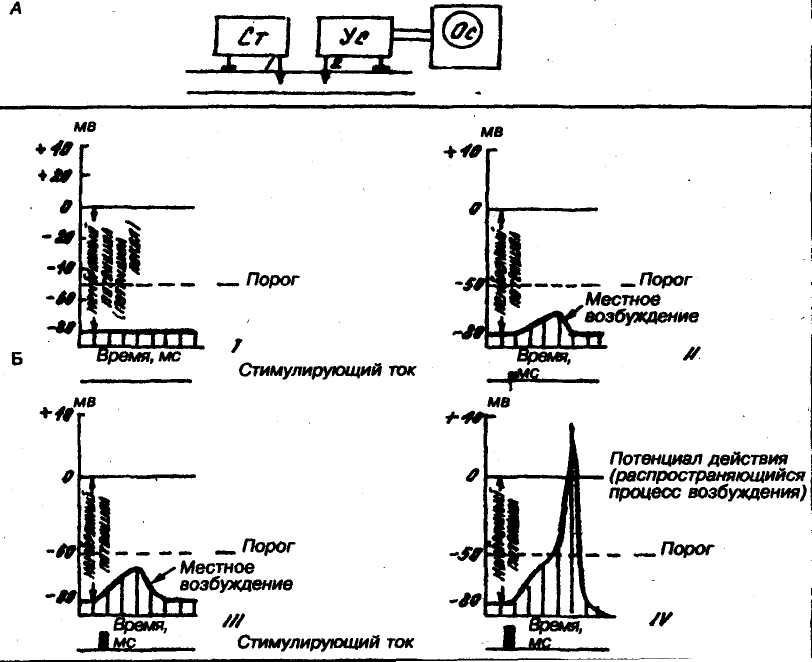
Рис.
3. Изменю!
we
мембранного потенциала
и формирование потенциала действия
в зависимости от силы
раздражения.
А — схема опыта. Ст — стимулятор, Ус — усилитель, Ос — осциллоскоп; 1 — раздражающий электрод, 2 — отводящий электрод.
Б — графики регистрации потенциалов (схематические). I — мембранный потенциал, II — местное возбуждение, вызываемое слабым раздражением, 111 — местное возбуждение, вызываемое раздражением большей силы, (V — бегущая волна возбуждения при пороговом раздражении, с типичным «стойком».
и сследователь
получает возможность оценивать
возбудимость объекта и сравнивать его
с другими
возбудимыми объектами или оценивать
изменение возбудимостн во времени,
например, при
оценке длительности абсолютной
рефрактерной фазы.
сследователь
получает возможность оценивать
возбудимость объекта и сравнивать его
с другими
возбудимыми объектами или оценивать
изменение возбудимостн во времени,
например, при
оценке длительности абсолютной
рефрактерной фазы.
В данном (приведенном выше) примере с десятью аксонами мы можем сказать, что самый возбудимый аксон — это аксон под номером 1, а самая низкая возбудимость у аксонов под номерами 9 и 10.
Закон времени (или зависимость пороговой силы раздражителя от временя его действия). Этот закон утверждает: раздражитель, вызывающий возбуждение, должен быть достаточно длительным, воздействовать на ткань некоторое время, чтобы вызвать возбуждение. Оказалось, что в определенном диапазоне зависимость пороговой силы раздражителя от длительности его действия носит характер обратной зависимости (гипербола) — чем меньше по времени действует на ткань раздражитель, тем выше требуется его сила для инициации возбуждения. На кривой (Гоорвега-ВеЙса-Лапика) выделяют области, которые свидетельствуют о том, что если раздражитель достаточно длительный, то пороговая сила раздражителя не зависит от его длительности. Эта минимальная сила получила название «ре-
14
например, в клинике нервных болезней,
в хирургии при лечении ранений нервов.
Возбудимые ткани
существенно отличаются друг от друга
по этим показателям. Например, у нервов,
снабжающих переднюю группу проксимальных
мышц верхних конечностей хронаксия
равна 0,08—0,16 мс, а
у мышц — 0,2—0,5 мс, т. е. намного больше.
При поражении нерва хронаксия
увеличивается. В физиологии и клинической
практике используется специальный
прибор — хронаксиметр, позволяющий
определить хронаксию и реобазу мышц
(двигательная
хронаксия), чувствительных нервных
волокон (чувствительная хронаксия),
вестибулярного
аппарата (при раздражении сосцевидного*
отростка), сетчатки (вспышки света при
ее электрической стимуляции).
Второе важное
следствие этого закона: слишком короткие
по длительности импульсы не способны
вызвать возбуждение, каким бы сильным
ни был стимул. Это применяется в
физиотерапии:
используют токи высокой частоты для
получения,калорического эффекта..
Закон градиента.
Для того, чтобы раздражитель вызвал
возбуждение, он должен нарастать
достаточно быстро. Если раздражитель
нарастает медленно, то в силу развития
аккомодации (инактивации натриевых
каналов), происходит повышение порога
раздражения, поэтому
для получения возбуждения величина
стимула должна быть больше, чем если
бы он нарастал мгновенно. Зависимость
величины пороговой силы раздражителя
от скорости его
нарастания тоже носит гиперболический
характер (является обратно-пропорциональной
зависимостью). Минимальный градиент —
это минимальная скорость нарастания
раздражителя,
при которой ткань еще способна ответить
возбуждением на данный раздражитель.
Этот показатель тоже используется для
характеристики возбудимости. Нерв,
обладающий более высокой
возбудимостью, чем скелетная мышца,
быстрее аккомодирует, поэтому
минимальный градиент у него выше
(например, 10 мА/с), чем у мышцы (2 мА/с, к
примеру). В
практике, исходя из существования
такого закона, для нанесения электрического
15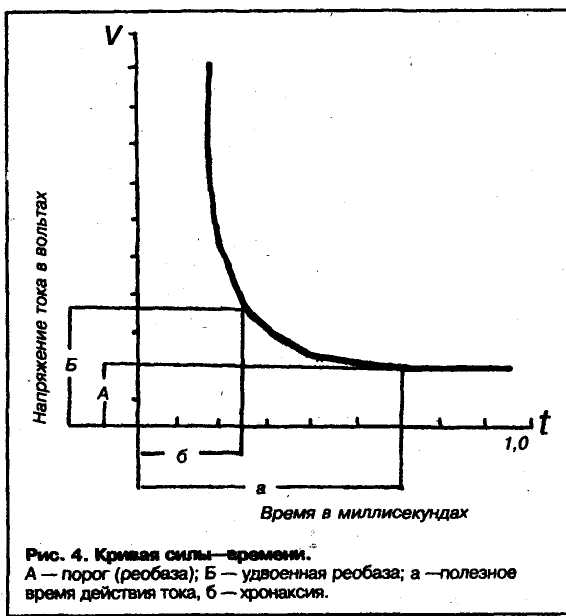
обаза». Начиная с некоторой величины длительности импульса, пороговая сила его зависит от длительности ■— чем меньше длительность, тем выше должна быть сила раздражителя. Вводится понятие «полезное время» — минимальное время, в течение которого раздражитель панной силы должен воздействовать на ткань, чтобы вызвать возбуждение. Если сила раздражителя равна двум реобазам, то полезное время для такого раздражителя получает еще одно название — хро-каксия. (Итак, хронаксия .— это полезное время раздражителя, сила которого равна 2 реобазам). В клинической медицине и в физиологии реобаза и хронаксия широко применяются для оценки состояния возбудимых тканей,
раздражения на возбудимую ткань с целью оценки ее функционального состояния обычно используют прямоугольные электростимулы — стимулы, у которых фронт нарастания очень высокий (бесконечно быстрый). Для определения минимального градиента и других показателей, характеризующих свойство аккомодации, используются пилообразные токи; наклон пилы регулируется, и это позволяет определить минимальный градиент.
В целом, закон градиента имеет и другие аспекты» например, методика применения лекарственных веществ, закаливающих процедур.
СООТНОШЕНИЕ ФАЗ ПОТЕНЦИАЛА ДЕЙСТВИЯ И ВОЗБУДИМОСТИ
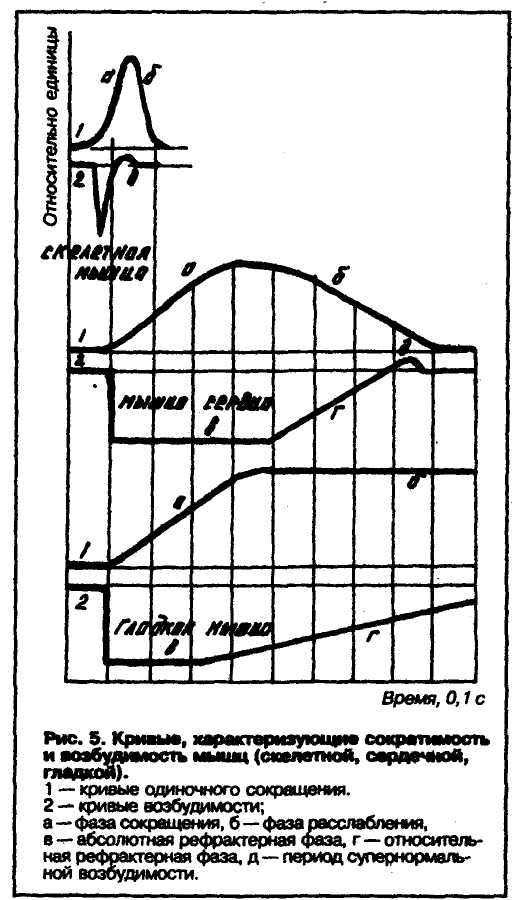 Когда
ткань возбуждается — генерирует ПД, то
временно (соответственно с длительностью
ПД) в ней меняется возбудимость: вначале
ткань становится совершенно невозбудимой
(абсолютная рефрактерность) — любой по
силе стимул не способен вызвать в ней
новый приступ
возбуждения. Эта фаза
обычно наблюдается во время
пика ПД. Затем происходит постепенное
восстановление возбудимости до
исходного состояния (фаза
относительной рефрактерно-сти)
— в этот момент раздражитель
может вызвать возбуждение (генерацию
нового ПД), но для этого
он должен быть намного больше
порогового (исходного). Затем
(в фазу следовой негативности)
возбудимость повышается (супервозбудимость,
или фаза экзальтации).
В этот момент подпорого-вые
раздражители могут вызывать возбуждение.
Наконец, в тканях, в которых
ярко проявляется следовая
гиперполяризация, наблюдается
еще одна фаза — субнормальной
возбудимости (сниженной возбудимости).
Когда
ткань возбуждается — генерирует ПД, то
временно (соответственно с длительностью
ПД) в ней меняется возбудимость: вначале
ткань становится совершенно невозбудимой
(абсолютная рефрактерность) — любой по
силе стимул не способен вызвать в ней
новый приступ
возбуждения. Эта фаза
обычно наблюдается во время
пика ПД. Затем происходит постепенное
восстановление возбудимости до
исходного состояния (фаза
относительной рефрактерно-сти)
— в этот момент раздражитель
может вызвать возбуждение (генерацию
нового ПД), но для этого
он должен быть намного больше
порогового (исходного). Затем
(в фазу следовой негативности)
возбудимость повышается (супервозбудимость,
или фаза экзальтации).
В этот момент подпорого-вые
раздражители могут вызывать возбуждение.
Наконец, в тканях, в которых
ярко проявляется следовая
гиперполяризация, наблюдается
еще одна фаза — субнормальной
возбудимости (сниженной возбудимости).
У аксона, у сомы нервной клетки, у скелетных мышц длительность абсолютной рефрактернос-ти составляет 0,5—2 мс, длительность относительной рефрактер-ностн — 5—10 мс, длительность супернормальной возбудимости — в пределах 20—50 мс. У сердечной мышцы длительность АРФ около 300 мс — намного больше и это имеет большой физиологический смысл — это период, запрещенный для нового сокращения. В основе рефрактерности лежат процессы инактивации натриевых каналов и восстановления активности этих каналов.
16
Как определить длительность АРФ, ОРФ? Для этого обычно объект исследования раздражают двумя стимулами: вначале наносится 1-й стимул (надпороговый), в ответ на который ткань генерирует ПД, затем, с регулируемым (по желанию экспериментатора) интервалом подается 2-й стимул той же силы. Если он вызывает ПД, то исследование повторяют и интервал между 1-м и 2-м стимулом уменьшают. Эта процедура длится до тех пор, пока второй стимул не будет способен вызвать ПД, даже если величина стимула будет намного превышать исходное значение.
МЕТОДЫ ИССЛЕДОВАНИЯ ВОЗБУДИМОСТИ НЕРВОВ И МЫШЦ
Для исследования возбудимости можно применить такие показатели как порог раздражения, реобазу, хронаксию, скорость аккомодации, минимальный градиент, длительность АРФ и ОРФ (см. выше). Лабильность возбудимых тканей определяется по максимальному числу возбуждений в единицу времени в ответ на ритмические раздражения, а также по длительности абсолютной рефрактерной фазы.
ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ТКАНИ
В 1859 г. Пфлюгер сформулировал закон полярного действия тока: при действии постоянного тока возбуждение происходит только в момент замыкания или только в момент размыкания цепи постоянного тока. При этом в момент замыкания оно возникает под катодом, а в момент размыкания — под анодом. Почему важен этот закон и сейчас, более 100 лет с момента его открытия? Дело в том, что понимание этого закона дало возможность объяснить возбуждение и процессы, лежащие в его основе. Он объясняет механизмы, лежащие в основе изменения КУД. Итак, этот закон — пробный камень общей физиологии возбудимых тканей.
При замыкании цепи постоянного тока изменяется сила раздражителя. Поэтому возбуждение возникает либо в момент замыкания, либо в момент размыкания (по сути — это закон градиента). При замыкании под катодом происходит деполяризация, и если она достаточна для достижения КУД, то возникает возбуждение — генерируется ПД. Если этого не происходит, то в области катода изменяется возбудимость: она возрастает вначале (эле-ктротон, катэлектротон), но потом, вследствие инактивации натриевых каналов резко снижается. Это получило название католической депрессии. Явление депрессии открыл пермский физиолог Б. Ф. Вериго. Это явление, вероятно, лежит в основе такого процесса, как пресинаптическое торможение. Под анодом при замыкании цепи постоянного тока повышается величина мембранного потенциала — возникает гиперполяризация. Поэтому при замыкании цепи тока возбуждение не возникает. Возбудимость снижается (явление анэлек-трона). Однако, если ток Действует долго, то в условиях гиперполяризации КУД снижается (МП приближается к исходному уровню), поэтому при размыкании цепи тока в области анода возникает возбуждение (быстро уменьшающаяся величина мембранного потенциала «достигает» КУД).
Итак, явление анэлектротона и феномен появления возбуждения в области анода при размыкании цепи постоянного тока вскрыли важный механизм — изменение КУД в процессе функционирования возбудимых тканей.
Закон полярного действия тока может использоваться и. на практике: если требуется заблокировать проведение возбуждения по нерву (болевая рецепция, например), то можно использовать постоянный ток, при этом в области расположения анода возбудимость будет снижена, что приведет к блоку проведения возбуждения. Аналогично — при длительной деполяризации в области катода (когда возникает католическая депрессия).
2. Физиология человека