

2. Межвидовая конкуренция. Логистическая модель межвидовой конкуренции.
Об интенсивности межвидовой конкуренции судят обычно по степени подавления скорости роста каждой популяции или же по изменению состояния особей, образующих данные популяций (например, по уменьшению их размеров). Не следует забывать и о том, что скорость роста популяции - это интегрирующий показатель, вместо которого можно использовать и отдельные его составляющие, т.е. рождаемость и смертность. Очевидно, что влияние конкуренции может проявляться в снижении рождаемости и(или) в возрастании смертности.
В отличии от внутривидовой конкуренции, межвидовая конкуренция в разных частях ареала и(или) в разные моменты времени она может сильно варьироваться (как по природе оспариваемого ресурса, так и по механизмам взаимодействия), поскольку в каждом конкретном случае представители данного вида могут вступать в конкуренцию с разными видами. Именно поэтому возможность коэволюции конкурирующих видов вызывает среди экологов споры. Очевидно, что если какой-то вид конкурирует с другими в процессе своей эволюции, то выжить он может, либо избегая конкуренции (например, перейдя на другой ресурс или мигрируя в другое местообитание), либо развивая свою конкурентоспособность и усиливая давление на конкурентов.
Межвидовую конкуренцию, так же как и внутривидовую можно разделить на 2 основных типа - интерференцию и эксплуатацию, хотя в любом конкретном взаимодействии можно выявить элементы и того, и другого типа. В случае эксплуатационной конкуренции особи взаимодействуют друг с другом косвенно, реагируя на количество ресурса, пониженное вследствие активности конкурентов. Пример такого взаимодействия дают эксперименты Тилмана и его коллег на диатомовых водорослях (рис. 7). Когда межвидовая конкуренция основана на использовании общего ресурса, то один вид потребляет этот ресурс и уменьшает его количество до такого уровня, при котором скорость роста, размножения или выживаемость другого вида снижается.
Морские желуди, которых изучал Коннелл, являются примером интерференционной конкуренции (рис. 8). Balanus, в частности, прямо и физически препятствует закреплению Chthamalus на ограниченных участках каменистого субстрата.
Другим важным моментом является то, что межвидовая конкуренция (подобно внутривидовой) часто бывает сильно асимметричной, что предполагает не одинаковы последствия для обоих видов. Например, в случае, исследованном Коннеллом, Balanus вытеснял Chthamalis из зоны возможного сосуществования; но любое воздействие Chthamalis на Balanus было незначительным (рис. 8).
Наконец, конкурирующим за один ресурс видам часто приходится использовать другой лимитированный ресурс.

Рис. 7. Результаты экспериментов по раздельному (А, Б) и совместному (В) культивированию двух видов диатомовых водорослей.
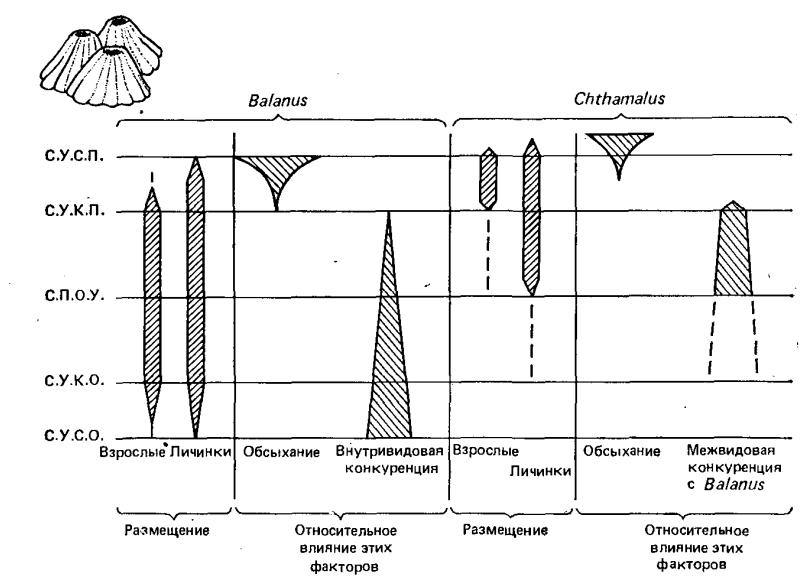
Рис. 8. Размещение взрослых особей и вновь осевших личинок и в приливно-лтливной зоне. Слева показаны средние уровни: СУСП –сизигинного прилива, СУКП – квадратурного прилива, СПОУ – приливно-отливный, СУКО – квадратурного отлива, СУСО – сизигийного отлива.
Первые математические модели конкуренции были предложены в конце 20-х гг. В. Вольтеррой, а несколько позднее независимо от него А. Лоткой. Предпосылки, лежащие в основе моделей Вольтерры и Лотки, очень близки, как и сами дифференциальные уравнения, описывающие изменения численности конкурирующих видов. Согласно В. Вольтерре, скорости роста популяций двух видов, потребляющих одну и ту же пищу, можно записать следующими уравнениями:
dN1/dt = [ε1 – γ1F(N1N2)]N1,
dN2/dt = [ε2 – γ2F(N1N2)]N2,
где N1 и N2 - численности 1-го и 2-го видов, а выражения в квадратных скобках являются коэффициентыми прироста популяций, предоставленные как разности между показателями, характеризующими рост популяций при отсутствии лимитирования по пище (ε1 и ε2 соответственно для 1-го и 2-го видов), и показателями, характеризующими ограничение роста недостатком пищи γ1F(N1N2) для 1-го вида и γ2F(N1N2) - для 2-го. Ограничение роста популяции, вызванное увеличением собственной численности и численности конкурента, для обоих видов описывается в уравнениях Вольтерры одной и той же функцией F(N1N2), а что касатся коэффициентов γ1 и γ2, то они характеризуют только количественное выражение (интенсивность) этого процесса.
Эти уравнения довольно трудно интерпретируются в обычных понятиях, используемых биологами при анализе роста популяций. Поэтому в экологической литературе значительно большую популярность получила модификация данных уравнений, предложенная в 30-х гг. Г.Ф. Гаузе.
Данная модификация уравнений Вольтерры-Лотки базируется на следующих допущениях:
1) рост численности описывается логистической кривой. Следовательно в ней должны соблюдаться все исходные предпосылки логистической модели, а именно:
- линейное снижение удельной скорости популяционного роста по мере роста ее численности,
- наличие предельной численности К, при которой скорость прироста численности становится равной нулю,
- отсутствие каких-либо временных задержек (лаг-эффектов) в реакции скорости роста популяции на изменение численности.
2) тормозящее влияние, оказываемое популяцией одного вида на рост другого, также описывается линейной зависимостью. Соответственно коэффициенты конкуренции предполагаются постоянными величинами, а изоклины, отвечающие стационарному состоянию популяции, - прямыми.
Согласно уравнению Вольтеры-Лотки-Гаузе динамика численности первого вида описывается уравнением
dN1/dt = r1N1[(K1 – N1 – α12N2)/K1],
а 2-го вида
dN2/dt = r2N2[(K2 – N2 – α21N1)/K2],
где N1 и N2 - численности 1-го и 2-го видов в рассматриваемый момент времени t, r1 и r2 - показатели максимальной скорости их роста при значениях плотности, близких к нулю, a К1 и К2 - асимптоты логистического роста каждой из популяций в отсутствие конкурентов. Коэффициент α12 оценивает степень влияния 2-го вида на рост численности 1-го вида. Или, иначе говоря, коэффициент α12 показывает, во сколько раз тормозящее действие, оказываемое на рост популяции 1-го вида особью 2-го вида, отличается от аналогичного действия, оказываемого особью собственного (т.е. в данном случае 1-го) вида. Если α12 = l, то скорость роста популяции 1-го вида реагирует на увеличение численности 2-го вида совершенно так же, как на увеличение собственной численности. Аналогична образом коэффициент α21 оценивает тормозящее воздействие 1-го вида на рост 2-го в сравнении с воздействием особей 2-го вида. В случае, когда α12 = α21, конкуренты угнетают друг друга в равной степени.
Хотя уравнения и не решаются, их можно представить в графической форме, во всяком случае для популяций стационарных, т.е. не меняющих свою численность (рис. 9). Изменения численности 1-го вида в любой точке данного поля координат будем изображать горизонтальным вектором, а численностт 2-го вида - вертикальным.
Совмещение графиков изоклин 1-го и 2-го видов на одном координатном поле дает четыре варианта взаимного расположения изоклин (рис. 10).
Если изоклины не пересекаются, то выигрывает тот вид, изоклина которого идет дальше от начала координат, например, в случае (а) 1-й вид вытесняет 2-й благодаря своей способности увеличивать численность после того, как уже достигнута предельная численность 2-го вида; в случае (б) 1-й вид вытесняется 2-м.
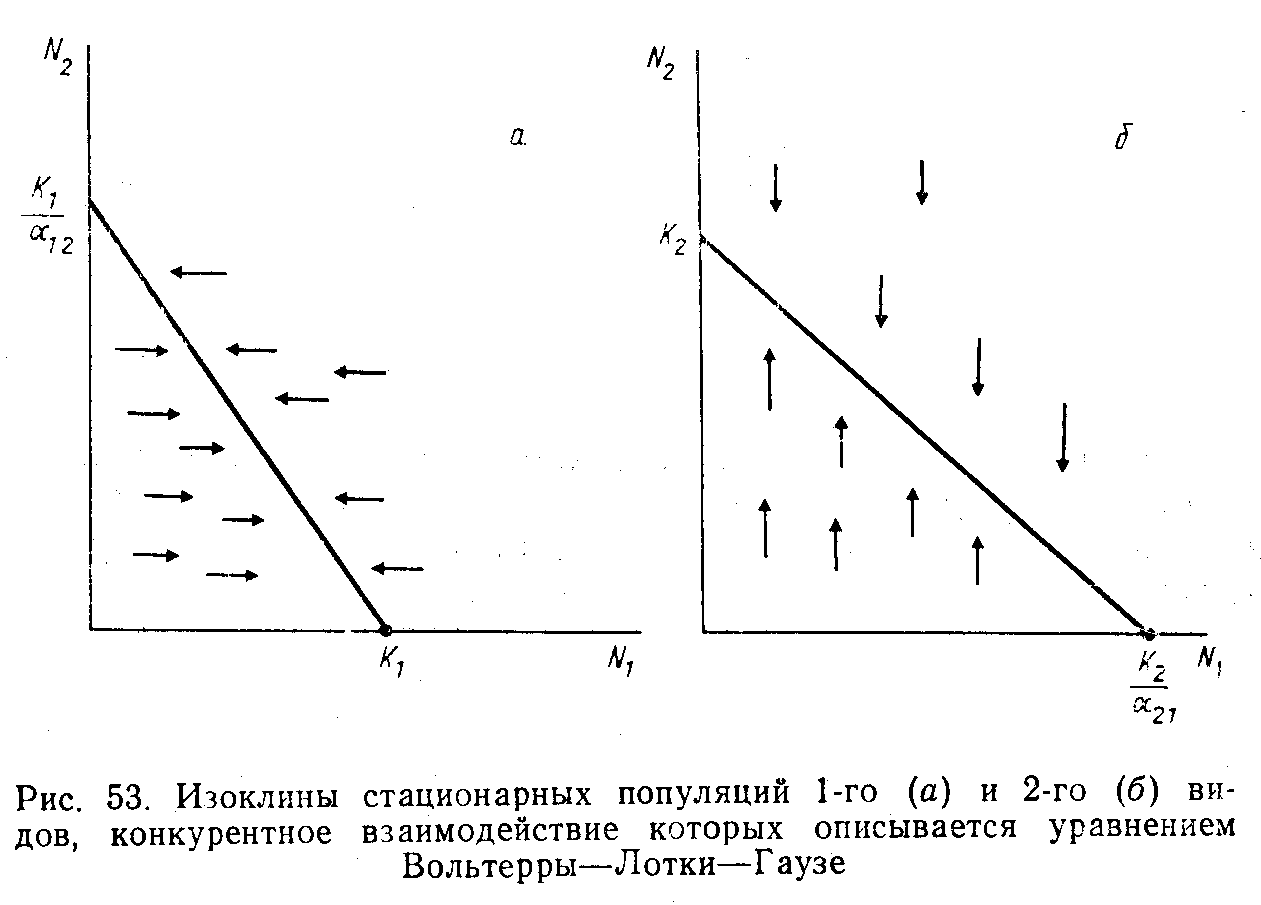
Рис. 9. Изоклины стационарных популяций первого (а) и второго (б) конкурирующих видов.
При пересечении изоклин возможно либо стабильное равновесие, когда векторы изменения численности направлены к точке пересечения (в), либо нестабильное равновесие, когда векторы направлены от точки пересечения (г). В последнем случае сосуществование практически не наблюдается, а исход конкуренции целиком определяется начальным соотношением численностей рассматриваемых видов. Таким образом, в трех из четырех теоретически возможных вариантов конкурентная борьба заканчивается элиминацией одного вида. Случай сосуществования соответствует ситуации, когда рост популяции каждого вида в гораздо большей степени зависит от собственной численности, чем от численности конкурирующего вида. Значения как α12, так и α21 должны быть при этом обязательно меньше единицы.
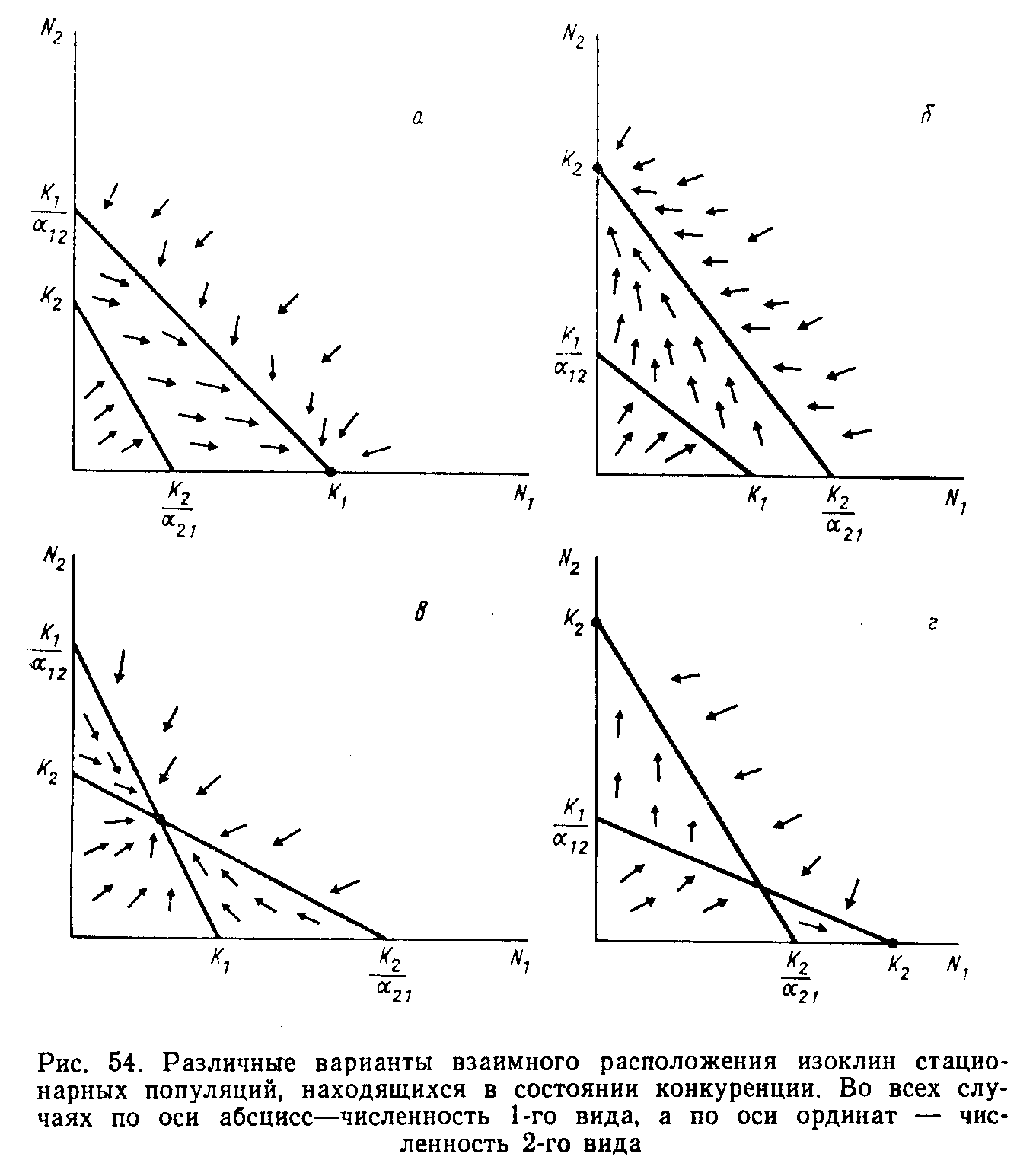
Рис. 10. Варианты взаимного расположения изоклин стационарных конкурирующих популяций.
Надо сказать, что сомнения относительно неизменности коэффициентов α12 и α21 в ходе роста численности конкурирующих популяций возникли уже при первых попытках экспериментальной проверки обсуждаемой модели. Так, Г.Ф. Гаузе изучая конкуренцию между разными видами инфузорий, показал, что коэффициенты α12 и α21 могут даже менять свой знак по мере роста популяций. В частности, в первые дни совместного культивирования Paramecium aurelia и Paramecium caudatum коэффициент α12, показывающий влияние Р. aurelia на Р. caudatum,, близок к —1, т.е. вместо —α12N1 в уравнении получаем +α12N1. Иными словами, присутствие Р. aurelia способствовало росту популяции Р. caudatum. Г.Ф. Гаузе поясняет, что этот неожиданный эффект, по-видимому, связан с установлением более оптимального для Р, caudatum соотношения между плотностью используемых в качестве корма бактерий и плотностью самих инфузорий. При дальнейшем росте обоих видов стимулирующее действие Р. aurelia на Р. caudatum сменяется угнетающим, что отражается увеличением коэффициента α21, который становится равным +0,61.