

Chapter 22
Notch
Notched wings, Morgan, and the gene theory
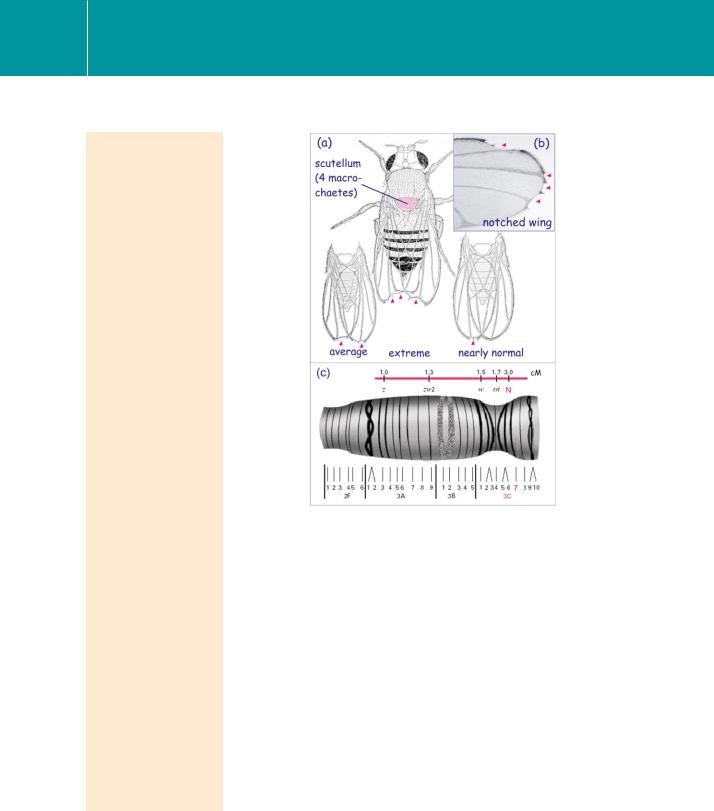
Notches in the wings of Drosophila were first described in 1916 by T. H. Morgan, who ascribed them to a spontaneous genetic mutation (Figure
22.1). Genes, the elements of heredity postulated by Gregor Mendel some 50 years previously, remained just that: postulates. Morgan appreciated that
development and heredity are inextricably linked and that knowledge in this area could not be advanced without an understanding of the nature of the gene. It fell to him to tackle the central question and introduce the science of genetics into embryology, or, as he phrased it, ‘approaching a physiological problem with a new and strange discipline to the classical physiology of
the schools’. This, and perhaps the fact that they were cheap to work with, explains his involvement with flies.
In writing these paragraphs, we have relied heavily on the work of Scott Gilbert, see http://8e.devbio.com. Details can be found in Gilbert.1,2
699
Signal Transduction
Thomas Hunt Morgan was awarded the Nobel Prize in Physiology or Medicine in 1933 ‘for his discoveries concerning the role played by the chromosome in heredity’.
‘A defect in the protoplasm often brings about a modified cleavage and also a defective embryo, and this takes place even though the whole of the nuclear material
of the unsegmented egg remains present. There seems, therefore no escape from the conclusion that in the protoplasm, and not in the nucleus, lies the differentiating power of the early stages of development’.
‘Insofar as it carries a nucleus, every cell, during ontogenesis, carries the totality of all primordia; insofar as it contains a specific cytoplasmic cell body, it is specifically enabled by this to respond to specific effects only … When nuclear material is activated, then, under its guidance, the cytoplasm of the cell that had first influenced the nucleus is in turn itself changed, and thus the basis is established for a new elementary process, which itself is not only a result but also a cause’.
FIG 22.1 Appearance of the Notch phenotype.
(a) Notched wings, first described by Morgan and drawn by Edith Wallace, artist and curator of the Drosophila stocks at Columbia University. (b) A photograph of a notched wing. Note the intermittent absence of small sensory bristles (microchaete) along the edge. (c) Localization of the Notch locus on the polytene salivary X chromosome of Drosophila melanogaster. z, zeste; zw2, zeste-white2; w, white; N, notch; rst, roughest. Locus distance in centimorgans (cM). 1 cM represents 300 kilobases.
Image (a) from Morgan.3 Image (b) from ‘The interactive fly’, Thomas B. Brody (www.sdbonline.org), section ‘Genes involved in tissue and organ development’.
At that time there were two dominant ideas concerning the question of what determines embryonic development. Some declared that the answer must lie in the distribution of the protoplasm. Certain cells contained, the material required to constitute individual organs, while others lacked it. Others
had it that all the information resides in the nucleus. Thus, different sets of information would be retained in some cells, not in others, in this way guiding the stages of development.
Initially Morgan denied the evidence for the ‘nuclear preformation’ theory. This belief was founded, in part, on his own experiments, which revealed
700

Notch
the impotence of the nucleus when a segment of cytoplasm was removed. Hans Driesch eventually revealed that the two opposing theories
– progressive determination of the cytoplasm by the nucleus versus cytoplasmic control of nuclear function – are like the two sides of a coin. Working with sea urchin eggs, he showed that the nucleus of a cell that is destined originally to become ectoderm can retain the ability to become endoderm or even to generate the entire embryo itself. Driesch correctly concluded that nucleus and cytoplasm interact to guide all the succeeding stages of development.
Compelling evidence for nuclear control of phenotype came from experiments designed to resolve the mechanism of sex determination. Studies of male gametogenesis in the lubber grasshopper, by Sutton and McClung, demonstrated that half of the spermatocytes carry an ‘accessory chromosome’ that has an effect on sex determination following fertilization. Henking had already noted this accessory chromosome in plants, and not knowing its function, labelled it ‘X’, henceforth the identifier of the larger
of the two universal sex chromosomes. Further evidence came from Nettie Stevens, who studied the chromosomes of mealworm beetles. She reported a
. . . clear case of sex determination, not by an accessory chromosome but by a definite difference in the character of the elements of one pair of chromosomes of the spermatocyte of the first order, the spermatozoon which contains the small chromosome determining the male sex.
In order to resolve this conflict Morgan commenced an investigation of sex-limited inheritance in Drosophila. In 1910, assisted by some whiteeyed mutant flies that had entered his stocks by chance, he found that the inheritance of red-eyes is inseparable from the X chromosome. Indeed, the factors that determine eye colour, body colour, wing shape, and sex all
segregate together with the X chromosome. This constituted direct evidence for the determination of sex by the chromosomes: he had the provided evidence that genes are heritable elements localized on chromosomes and that their absence or presence affects the phenotype. This finding is known as the gene theory.
In 1913, Morgan and A. H. Sturtevant presented the first physical genetic map of the X chromosome of Drosophila. From a continuous search of mutants and crossing experiments, genetic maps of other chromosomes came to light. The notched-wing Drosophila played a part in all this. By 1933, developmental biology had advanced, especially with the findings of Spemann and others (see page 618). It was understood that environmental factors, operating by cell-to-cell contact or diffusible factors, influence cell fate (the discovery of the ‘organizer’). But again the problem of how these interactions change the phenotype of cells remained unsolved.
‘From the point of view under consideration, results of this kind are of interest because they bring up once more, in a
slightly different form, the problem as to whether the organizer acts first on the protoplasm of
the neighboring region with which it comes in contact, and through the protoplasm of the cells on the genes; or whether the influence is more directly on the genes. In either case the
problem under discussion remains exactly where it was before. The evidence from the organizer
has not as yet helped to solve the more fundamental relation between genes and differentiation, although it certainly marks an important step forward in our understanding of embryonic development’.4
701

Signal Transduction
We now know that cell environmental factors, acting through receptors, operate signal transduction pathways that directly or indirectly (by changes in protein translation) affect the activity of factors that lead to induction or suppression of transcription. The Notch signal transduction pathway is no exception. It modifies cell fate by silencing some genes and inducing the expression of others, mediating an essential and perhaps universal function in the assignation of cell fates during development.5 Notch is just one example of how environment, in this case the neighbouring cell, influences cell fate, but its mode of action provides a perfect and even simple answer to the question that Morgan had been asking throughout his career.
Unlike any other system that we have discussed, Notch acts through ligandinduced cleavage of its own intracellular segment that moves straight to the nucleus where it regulates transcription.
One gene, many alleles
The Notched wing Drosophila is the consequence of null alleles, abnormal genes that result in loss of function of the gene product. They are the consequence of nonsense mutations, a translocation or an inversion of the chromosome. Null alleles for Notch are lethal in hemizygous males (XY) and only viable in heterozygous females in which they are partially compensated by a functional gene. There are numerous lethal Notch alleles, all indicated as N with superscripts: N 40, NNIC, etc. Females carrying one mutated N allele
display the characteristic notched wings, thickened wing veins, and some bristle abnormalities. There are many other, non-lethal, Notch alleles, the consequences of a partial loss of function due to missense mutations, partial deletions, or very low levels of expression. One group, the‘recessive visibles’, which fall in three complementation groups, facet (fa), split (spl), and notchoid (Nd), affect either wing or eye morphology. Yet another class comprises the dominant Abruptex (Ax) mutations, which cause missense mutations in the extracellular (ligandbinding) domain of Notch and, surprisingly, enhance the Notch signal.
When Poulson commenced his analysis of flies that had lost the whole or certain bands of the X chromosome, including the band that contains the Notch locus, he discovered a bewildering array of morphological
abnormalities affecting nearly all parts of the body. An excess of neural tissue was particularly apparent. This provided the first evidence that Notch has
an important role in the early stages of development.6,7 This was confirmed and extended by the work of Campos-Ortega8 who demonstrated that the neural hypertrophy is due to a defect that arises very early, roughly 4 h after fertilization. At this stage, precursor cells (neuroblasts) segregate from others that constitute the neurogenic ectoderm. Normally, only the neuroblasts give rise to neural tissue, the remainder of the neurogenic ectoderm becoming epidermis.9 However, when Notch is absent, nearly all cells proceed towards
702