


И.Л. ДИКИЙ И.Ю.ХОЛУПЯК Н.Е.ШЕВЕЛЕВА М.Ю.СТЕГНИЙ
МИКРО БИОЛОГИЯ
Для студентов фармацевтических вузов и фармацевтических факультетов медицинских институтов
Под редакцией д-ра мед. наук, проф., акад. АНТК Украины, И. Л. ДИКОГО
Харьков «Прапор»
Издательство УкрфА 1999
ББК 52.64
Д45
Рекомендовано Науково-методичнию komichw аз фармацию
Mimіcmepcmea oceimu Украни
як піручник для студентів фармацевтичних вузів
та фармацевтичних факультетів медичних інcmumymiв
Рецензенти:
А. Я. Циганенко, професор,
зав. кафедрою мжробюлоги, вірусології та імунології
Харківського державного медичного університету,
академік вищої школи Украни
А. А. Воробйов, професор, зав. кафедрою
мікробіології з вipyсoлогією та імунологіею
Московської медичної академії ім. І. М. Сеченова,
академік РАМН, академік міжнародної академії наук
ВИЩ0ї ШКОЛИ
В учебнике обобщены современные достижения в области медицинской, фармацевтической, санитарной и химической микробиологии. Представлены фундаментально-прикладные положения по морфологии, физиологии прокариотов, эукариотов и вирусов; рассмотрены методы культивирования бактерий и вирусов, механизмы взаимодействия вирусов с клеткой. Освещены вопросы инфекции, химиотерапии, иммунитета, иммунопатологических состояний, иммунобиотехнологии. Обобщены материалы по микроэкологии, основам асептики и антисептики, микробной контаминации лекарственного сырья и препаратов. Изложены сведения о возбудителях инфекционных болезней.
Рассчитан на студентов и преподавателей фармацевтических и медицинских высших учебных заведений.
Д
Без оголош.
218-99
ISBN5-7766-0755-8ISBN966-615-005-0
© І. Л. Дикий, І. Ю.
Холупяк, Н, Ю. Шевельова,
М. Ю. Стегній,
1999 © Видавництво «Прапор», 1999 ©
Видавництво У крФА, 1999
СПИСОК СОКРАЩЕНИЙ:
АІDS(от англ.acquiredimmunodeficiencysyndrome) — синдром приобретенного иммунодефицита человека (СПИД).
DDC— хивид, противовирусный препарат.
DDI— видекс, противовирусный препарат.
DLМ (от лат. dosisletalisminima)—наименьшее количество микробов, которое вызывает гибель 95 % зараженных восприимчивых лабораторных животных определенноговида и стандартной массы.
FAB-фрагмент молекулы Ат (иммуноглобулина) (от англ.fragmentantigenbinding) — фрагмент, связывающий антиген.
Fc-фрагмент молекулы Ат (иммуноглобулина) (от англ. fragment crystalline) — кристаллический фрагмент.
GMP(от англ.goodmanufacturingpractice) — надлежащая производственная практика (НПП), свод обязательных принципов, норм и правил в химико-фармацевтическом производстве.
НА — гемагглютинин, поверхностный антиген вириона гриппа.
HIV (от англ. human immunodeficiency virus) — вирус иммунодефицита человека (ВИЧ).
HLA(от англ.humanleucocyteantigenssystemA) — генетический комплекс главной системы гистосовместимости человека.
HTLV(от англ.humanT-lymphotropicvirus) — вирус человека,
обладающий тропизмом к Т-лимфоцитам. IHBcAg— сердцевинный антиген вируса гепатита В.
HBsAg— поверхностный антиген вируса гепатита В.
Ig
LAV
LD
50
минимальная доза микроорганизмов, вызывающая гибель 50 % экспериментальных животных.
NA— нейраминидаза.
NNN-среда — среда Нови — Николь — Ниль.
Аг — антиген.
Аг — Ат — иммунный комплекс антиген — антитело, образующийся в результате их специфического взаимодействия.
АДС-анатоксин — дифтерийно-столбнячный анатоксин.
АЗТ — азидотимидин, противовирусный препарат.
АКДС — ассоциированная коклюшно-дифтерийно-столбнячная вакцина.
АЛС — антилимфоцитарная сыворотка.
Ат — антитело (син. иммуноглобулина — Ig).
БКЯ — болезнь Крейцфельдта — Якоба.
БЦЖ (от фр. BacilledeCalmetteetdeGuerin—BCG) — живая противотуберкулезная вакцина, полученная французскими учеными А. Кальметтом и Ш. Гереном.
ВГА — вирус гепатита А.
ВИЧ — вирус иммунодефицита человека.
ВОЗ — Всемирная организация здравоохранения.
ГА — гепатит А.
ГГСИ — госпитальные гнойно-септические инфекции.
ГЗТ — гиперчувствительность замедленного типа.
ГЛС — готовые лекарственные средства.
ГНТ — гиперчувствительность немедленного типа.
ГСИ — гнойно-септические инфекции.
ГЦ-пары — гуанин-цитозин-пары.
ИД — иммунодиффузия.
ИФА — иммуноферментный анализ.
ИЭМ — иммунная электронная микроскопия.
ИЭФ — иммуноэлектрофорез.
КОКАВ — концентрированная культуральная антирабическая вакцина.
КУА — казеиново-угольный агар.
МКМ — микрометр (10-6м).
МНС (от англ. majorhistocompatibilitycomplex) — главный комплекс гистосовместимости, группа сцепленных генов, играющих главную роль в совместимости тканей при трансплантации.
МПА — мясо-пептонный агар.
МПБ — мясо-пептонный бульон.
МРВ-А— медленно реагирующие вещества при анафилаксии, жирные кислоты — медиаторы аллергии.
МФФ — Международная федерация фармацевтов.
НАГ-вибрионы — вибрионы, неагглютинирующиеся типовыми холерными сыворотками.
НМ - нанометр (10-9м).
ОКЗ — острые кишечные заболевания.
ОРЗ — острые респираторные заболевания.
ПАВ — поверхностно-активные вещества.
ПАСК — парааминосалициловая кислота.
PA— реакция агглютинации.
РБН — реакция биологической нейтрализации.
РИА (от англ. radioimmunoassay—RIA) — радиоиммунный
анализ.
РИАГА — реакция иммуноадгезивной гемагглютинации.
РИТ — реакция иммобилизации трепонем.
ЗШФ — реакция иммунофлюоресценции.
РМА — реакция микроагглютинации.
РН — реакция нейтрализации.
РНГА (РПГА) — реакция непрямой (пассивной) гемагглютинации.
РНП — рибонуклеопротеин.
РОЭ — реакция оседания эритроцитов.
РП — реакция преципитации.
РСК — реакция связывания комплемента.
РТ — ретикулярные тельца, вегетативные формы хламидий.
РТГА — реакция торможения гемагглютинации.
РТПХ — реакция «трансплантат против хозяина».
РХПТ — реакция «хозяин против трансплантата».
РЭМА — реакция энзимомеченых антител.
РЭС — ретикулоэндотелиальная система.
СИБ — система индикаторных бумажек, используемая для видовой дифференциации бактерий по ферментативным свойствам.
СКВ — системная красная волчанка.
СМЖ — спинномозговая жидкость.
СМФ — система мононуклеарных фагоцитов.
СОЭ — скорость оседания эритроцитов.
СПИД — синдром приобретенного иммунодефицита человека.
Т50- период времени, за который концентрация препарата в
крови уменьшается на 50 %.
ТМС - туберкулопротеидная маточная субстанция, белковая
фракция микобактерий туберкулеза.
ТЦД50/МЛ— тканевая цитопатическая доза вируса, вызывающая
поражение 50 % монослоя клеток.
УФ-лучи — ультрафиолетовые лучи.
ФГА - фитогемагглютинин, митоген растительного
происхождения.
ФПР - фрагмент К-плазмиды, ответственный за перенос ре-
зистентности.
ЦМ - цитоплазматическая мембрана. ЦНС - центральная нервная система.
ЦПД - цитопатическое действие (вирусов, токсинов и прочее)
в культуре клеток.
ЭИКП - энтероинвазивная кишечная палочка.
ЭПКП - энтеропатогенная кишечная палочка. элементарные ЭТ - тельца, спороподобные формы хламидий.
ЭТКП - энтеротоксигенная кишечная палочка.
ПРЕДМЕТ, ЦЕЛИ ЗАДАЧИ МИКРОБИОЛОГИИ
Микробиология (от греч. micros— малый,bios— жизнь,iogos— учение) — наука о мельчайших, невидимых невооруженным глазом микроорганизмах растительного или животного происхождения. Микробиология является одной из отраслей общей биологии и изучает закономерности жизни и развития Микроорганизмов в единстве с условиями среды их обитания,атакже тех изменений, которые они вызывают в организмах животных и растений. Эта наука разрабатывает методы использования микробов для нужд человека, пути и способы обезвреживания тех из них, которые являются опасными для жизни и здоровья людей.
Знание основ микробиологии крайне необходимо фармацевту в его профессиональной деятельности, поскольку без знаний в этой области немыслимо рациональное производство многих лечебно-профилактических средств: вакцин, сывороток, антибиотиков, гормонов и других препаратов, а также невозможно оценить порчу лекарственного сырья и разработать эффективные меры его предупреждения.
Особо следует отметить, что возникновение и быстрое развитие биотехнологии, приобретающей все большее значение в народном хозяйстве, базируется прежде всего на использовании микроорганизмов как продуцентов множества полезных веществ:
кормового белка, многих ферментов, антибиотиков, стероидных препаратов, аминокислот, витаминов и т. д. На использовании микроорганизмов основаны методы генной инженерии, позволяющие создавать новые штаммы с заданными биологическими свойствами.
Технология получения многих лекарственных средств требует асептических условий. Асептика и антисептика — это неотъемлемые компоненты знаний в области микробиологии, поэтому в каждой контрольно-аналитической лаборатории предусмотрен микробиологический контроль.
Провизору необходимы совершенно точные и конкретные знания о возбудителях заразных заболеваний человека и живот-
ных, лекарственных растений, способах передачи, патогенезе заболеваний, их лечении и профилактике.
Медицинскаямикробиология изучает морфологию, физиологию, отношение к внешним воздействиям среды, изменчивость микроорганизмов, основы инфекционного процесса, иммунитета, особенности возбудителей, патогенез инфекционных заболеваний, методы их лабораторной диагностики, специфического лечения и профилактики.
Кроме медицинской, микробиология имеет и ряд других самостоятельных научных разделов:
общая микробиологияизучает общие закономерности развития и жизнедеятельности микроорганизмов, их роль в круговороте веществ в природе, общие признаки наследственности и изменчивости;
ветеринарная микробиологияизучает болезнетворные микроорганизмы, вызывающие инфекционные заболевания у животных, разрабатывает методы их специфической диагностики, профилактики и лечения;
сельскохозяйственная микробиологияизучает роль микроорганизмов в почвообразовательных процессах, разрабатывает методы повышения плодородия почвы с помощью микроорганизмов, получения удобрений и кормов для животных;
пищевая микробиологияразрабатывает методы получения пищевых продуктов с помощью микроорганизмов, а также способы защиты пищевых продуктов от микробной порчи;
санитарная микробиологияизучает микрофлору воздуха, воды, почвы для гигиенической характеристики и оценки их как возможного источника и путей передачи инфекции; разрабатывает методы очистки воды, почвы, воздуха от микробов, прежде всего, патогенных;
техническая,илипромышленная микробиологияразрабатывает научные основы получения антибиотиков, витаминов, ферментов, пути и методы предохранения сырья от микробной порчи.
Фармацевтическая микробиология— это профессионально ориентированная дисциплина, интегрирующая основы медицинской, санитарной и промышленной микробиологии. Фармацевтическая микробиология изучает: возбудителей инфекционных заболеваний человека и растений; условия микробной порчи лекарственного сырья и контаминации препаратов в процессе изготовления и хранения; правила асептики, антисептики и дезинфекции при промышленном и экстемпоральном изготовлении фармацевтических препаратов; технологии получения антимикробных и иммунобиологических препаратов лечебно-профилактического и диагностического назначения.
ОСНОВНЫЕ ЭТАПЫ НАУЧНОГО СТАНОВЛЕНИЯ МИКРОБИОЛОГИИ
Медицинская микробиология является одной из фундаментальных дисциплин современного естествознания. «Медицина, которая служит человеку, слагается из науки и искусства, и над ними простирается чудесный покров героизма, без которого не может быть медицины». Это слова Гуго Глязера, автора широко известной книги «Драматическая медицина». Они по праву адресованы прежде всего микробиологам, гигиенистам и инфекционистам, доказавшим риском собственной жизни, всем историческим опытом борьбы с пандемиями и эпидемиями, что их главный жизненный девиз: «Светя другим, сгораю сам!».
На знамени драматической медицины героический поступок Макса Петтенкофера, выпившего в доказательство эпидемической природы холеры культуру возбудителя, и повторивших этот опыт И. И. Мечникова, Н. Ф. Гамалея, Д. К. Заболотного, И. Г. Савченко.
Первым привил себе чуму и погиб от нее английский врач
А. Уайт. Несмотря на трагический исход, последовали новые опыты по заражению себя чумой, их провели Р. Дженнет, А. Ф. Бюллар, А. Клот, А. Розенфельд, Э. Валли.
Высокую жертвенность во имя познания инфекционной сути желтой лихорадки проявили Н. Поттер, И. Л. Гюйон, К. Финлей, В. Рид, Д. Кэррол, Д. Ласеар.
История борьбы с малярией также чрезвычайно богата напряженными драматическими ситуациями, главными действующими лицами которой по праву признаны Р. Росс, Д. Грасси и А. Биньями. Последний сумел осуществить первое умышленное заражение малярией посредством укуса комара.
Еще до той поры, когда ученые приступили к изучению инфекционных болезней точными научными методами, в Одессе в 70-х годах XIX века русский врач О. О. Мочутковский провел на себе эксперимент с сыпным тифом. Пять раз он безрезультатно заражал себя и лишь на шестой — заболел сыпным тифом. Французский бактериолог Ш. Николь также благодаря опытам на себе установил, что промежуточным хозяином возбудителя сыпного
9
тифа является платяная вошь. За это открытие он в 1928 году был удостоен Нобелевской премии.
Русские ученые Г. Н. Минх и И. И. Мечников заражали себя возвратным тифом, Е. И. Марциновский (впоследствии директор Московского института тропических болезней) — лейшманиозом, итальянский врач Д. Профета — проказой. Эти опыты не были напрасными. Они помогли в борьбе против болезней, вызываемых бактериями и другими микроорганизмами. В конечном итоге эта борьба закончилась величайшими победами, вписавшими славные страницы в книгу прогресса медицины, в становление микробиологии как фундаментальной научной дисциплины.
Возникновение и развитие медицинской микробиологии как науки теснейшим образом связано с развитием знаний в изучении инфекционных болезней. Эпидемические болезни существовали на всех ступенях развития человеческого общества. Известно, что ликвидированная на планете в 1977 году натуральная оспа уносила миллионы жизней еще за 1120 лет до н. э. За 500 лет до н. э. было известно о бешенстве. Наблюдая за развитием эпидемий, люди пытались объяснить их происхождение. Гиппократ (460—370 гг. до н. э.) считал, что причиной эпидемии является вдыхаемый людьми воздух, который во время эпидемий содержит особые болезнетворные испарения — миазмы.
В 1546 году Д. Фракастро опубликовал трактат «О контагии, о контагиозных болезнях и лечении», в котором отмечается, что контагии представляют собой невидимые активные существа и заражение людей происходит в результате соприкосновения и даже на расстоянии. Это была гениальная догадка о существовании микробов.
Заслуга открытия микроорганизмов принадлежит А. ван Левенгуку (1632-1723), голландскому шлифовальщику стекол, сконструировавшему первый в мире микроскоп. При помощи этого инструмента, снабженного двояковыпуклой линзой с 300-кратным увеличением, Левенгук наблюдал клеточную структуру растений, зародышей, открыл существование сперматозоидов, а также различных микроорганизмов. В зубном налете, отварах и настоях он обнаружил невидимых невооруженным глазом «живых зверьков» — аnimalcula и зарисовал различные их формы — шаровидные, палочковидные, извитые. Левенгук — основоположник первого, морфологического периода в развитии микробиологии, давший последователям важнейшее инструментальное обеспечение — микроскоп. Однако сам Левенгук не мог указать, какую роль в природе и жизни человека играют открытые им микроорганизмы или, как назвал их в дальнейшем французский исследователь Седилло, микробы.
С открытия Левенгука начинается описательный период в развитии микробиологии. Спустя 100 лет О. Мюллер (1786) в Дании,
10
пользуясь более совершенным микроскопом, описал большое количество микроорганизмов, разделив их на два рода — Vibrio иMоnas. В 1838 году X. Эренберг на основании их признаков установил два семейства микроорганизмов Моnadina иVibrionia с родами Васterium,Spirillum,Sрirochaetа.
В 1775 году русский исследователь М. М. Тереховский впервые описывает экспериментальный метод исследования движения микроорганизмов, изучает влияние ряда физико-химических факторов (электрические заряды, вариации температуры, Кислотность и т. п.) на жизнеспособность микроорганизмов. Он описывает результаты своих весьма важных наблюдений относительно потребностей микроорганизмов в кислороде, впервые отмечает, что перед делением микроорганизмы растут и увеличиваются в размерах.
Однако в начале XIX века большинство исследований в области микробиологии носили описательный характер. Вторая половина XIX века ознаменовалась поистине революционными успехами в развитии медицинской микробиологии, чему в немалой степени способствовали достижения естествознания, такие как открытие клетки, закона превращения энергии, создание эволюционного учения Ч. Дарвина. Именно в это время жил и работал гениальный французский исследователь — химик и биолог по образованию — Луи Пастер. Роль этого выдающегося естествоиспытателя в бурном развитии микробиологии непереоценима. Вот как ее охарактеризовал В. Л. Омелянский: «С Пастера начинается второй, физиологический период в истории микробиологии... И если Левенгуку с полным основанием приписывается название «отца микрографии», то Пастер был истинным творцом и вдохновителем современной микробиологии, со сказочной быстротой разросшейся в огромную науку» (Избранные труды. Т. 2.— М., 1953.—С. 6).
Являясь основоположником изучения физиологии и биохимии микробов, Л. Пастер раскрывает сущность молочнокислого и маслянокислого брожения, показывает, что оно вызывается ни чем иным, как бактериями. Л. Пастер открывает анаэробный тип дыхания у бактерий, доказывает, что любое заразное заболевание имеет своего микроба-возбудителя.
Исследования в области брожения и самопроизвольного заражения послужили основой решения вопроса о «болезнях» вина и пива. В качестве меры борьбы Пастер предложил прогревать вино при 50—60º С. Эти исследования утвердили принципы стерилизации и метод пастеризации.
Основываясь на исследованиях Л. Пастера, Д. Листер в 1867 году ввел в хирургическую практику метод антисептики. В течение короткого времени Пастером были открыты возбудители гнойно-воспалительных заболеваний, куриной холеры и т. д. На примере
11
культуры возбудителя куриной холеры он впервые установил факт утраты ею вирулентности и способность этой ослабленной культуры создавать невосприимчивость к последующему заражению кур вирулентной культурой. Сделанный Пастером на основании этих опытов вывод о возможности применения ослабленных культур для формирования невосприимчивости к инфекционным заболеваниям создал предпосылки для развития нового научного направления — вакцинопрофилактики.
Доказывая правомерность своих выводов, Пастер получает сибиреязвенную, а затем антирабическую вакцины. Это было одно из величайших достижений в области медицины: открытие эффективного средства профилактического лечения болезни, которая в течение столетий считалась неизлечимой.
Второе, не менее славное имя в развитии микробиологии — Роберт Кох, который окончательно установил этиологию сибирской язвы (1876); разработал и применил плотные питательные среды, выделил чистые культуры; ввел в микробиологическую практику анилиновые красители, разработал метод иммерсии и микрофотографирования; обосновал и сформулировал триаду Генле — Коха, по которой признается роль микроба как возбудителя заболевания; заложил научные основы дезинфекции; открыл возбудителя туберкулеза и доказал инфекционную природу этого заболевания, получил туберкулин; открыл возбудителя холеры и изучил эпидемиологию этого заболевания.
Исключительно большое значение в развитии нового, оригинального направления микробиологии имеют исследования И. И. Мечникова — основателя учения об иммунитете — невосприимчивости организма к инфекционным заболеваниям. По определению Э. Ру, И. И. Мечников — это поэт микробиологии. Ему принадлежит приоритет творца фагоцитарной теории иммунитета, основоположника физиологического направления в иммунологии.
И. И. Мечников заложил основы учения об антагонизме бактерий, явившегося плодотворным источником для создания учения об антибиотиках, указал на огромное значение внешней среды для изменения свойств микробов, которые передаются по наследству, исследовал проблемы долголетия в связи с направленным изменением микрофлоры кишечника молочнокислыми бактериями. Совместно с Э. Ру он разработал модель экспериментального сифилиса, исследовал патогенез холеры. И. И. Мечников создал блестящую школу русских микробиологов, среди которых А. М. Безредка и Н. Ф. Гамалея, В. Л. Омелянский и И. Г. Савченко, Л. А. Тарасович и Д. К. Заболотный и многие другие.
В 1888 году французские ученые Э. Ру и А. Йерсен открыли дифтерийный токсин, а Ш. Китазато — столбнячный токсин.
12
В 1889—1892 годах Э. Беринг в Германии, Э. Ру во Франции, Я. Ю. Бартах в России независимо друг от друга получили антитоксическую дифтерийную сыворотку. В 1923 году Г. Рамон получил дифтерийный анатоксин. Эти два препарата позволили эффективно бороться с грозными, нередко смертельными инфекциями.
В конце XIX века, который по праву считается эрой бактериологии, было открыто огромное количество возбудителей инфекционных заболеваний:
1880 — возбудитель брюшного тифа (К. Эберт);
1883 — возбудитель дифтерии (Э. Клебс);
1891 — возбудители дизентерии (К. Шига);
1892 — вирус табачной мозаики (Д. И. Ивановский);
1894 — возбудитель чумы (А. Йерсен, Ш. Китазато).
В этот же период С. Н. Виноградский устанавливает роль микробов в круговороте азота в природе, вводит метод элективных питательных сред. В 1909 году Г. Риккетс открывает возбудителя сыпнотифозной лихорадки, а в 1913 — С. Провачек открывает возбудителя сыпного тифа.
В истории микробиологии начало XX века отмечено интенсивной разработкой проблемы специфической профилактики инфекционных заболеваний: вакцина против туберкулеза (А. Кальметт и Ш. Герен), противочумная вакцина (Г. Жирар и К. Робик), противотуляремийная вакцина (Б. Я. Эльберт, Н. А. Гайский), вакцина против полиомиелита (А. Сейбин, М. П. Чумаков, А. А. Смородинцев), дифтерийный и столбнячный анатоксины (Г. Рамон). В 1917 году д'Эррель открывает бактериофаги.
Этот период является основополагающим в развитии эффективных средств лечения инфекционных заболеваний — создается учение о химиотерапевтических препаратах и антибиотиках. Основатели химиотерапии — П. Эрлих, Д. Л. Романовский, Г. Домагк. Им принадлежит не только заслуга в обосновании основных требований к химическим препаратам, применяемым для лечения инфекционных заболеваний, но и обоснование условий их синтеза (метод химических вариаций П. Эрлиха), разработка химиотерапевтического индекса — основного показателя в характеристике этой группы препаратов, создание наиболее широко используемых в настоящее время сульфаниламидов.
С 1929 года благодаря исследованиям А. Флеминга по разработке пенициллина открывается новая эра в лечении инфекционных заболеваний — эра антибиотикотерапии. В развитии этого направления приняли участие Э. Б. Чейн и X. У. Флори, 3. В. Ермольева и С. Ваксман.
В 30-е годы нашего столетия в микробиологическую практику был введен электронный микроскоп, позволяющий детально
13
исследовать ультраструктуру вирусов и бактерий, разработаны основы микроскопической техники (ультратонкие срезы, гистохимические методы окраски).
ЭТАПЫ РАЗВИТИЯ ВИРУСОЛОГИИ
Честь открытия вирусов принадлежит Д. И. Ивановскому, который впервые в 1892 году на примере мозаичной болезни табака доказал существование новой формы возбудителя.
Слово «вирус» в древнеримском языке означало «яд», «отрава», оно применялось еще Л. Пастером для обозначения заразного начала.
В 1949 году происходит открытие, сыгравшее важную роль в истории вирусологии,— возможность культивирования клеток в искусственных условиях. В 1952 году Д. Эндерс, Т. Уэллер и Ф. Роббинс за разработку метода культивирования культуры клеток получили Нобелевскую премию. Появилась возможность выделения многочисленных новых вирусов, получения культу-ральных вакцин. Так, М. П. Чумаков и А. А. Смородинцев в содружестве с американскими вирусологами Дж. Солком и А. Сейбином разработали убитую и живую вакцины против полиомиелита. В 1959 году в нашей стране была проведена массовая иммунизация детей живой полиомиелитной вакциной, что позволило снизить заболеваемость и привело к практическому исчезновению паралитической формы болезни.
В современной вирусологии широко используют методы молекулярной биологии. Вирусы благодаря простоте их строения являются распространенной моделью для молекулярной биологии. Ни одно открытие молекулярной биологии не обходится без вирусной модели, включая генетический код, весь механизм внутриклеточной экспрессии генома. Развитие молекулярной биологии способствовало изучению первичной структуры нуклеиновых кислот и белков, появлению методов определения последовательностей аминокислот в структуре белка.
В 1972 году возникает новый раздел молекулярной биологии — генная инженерия. Появляется возможность получения большого количества нуклеиновых кислот и белков путем введения рекомбинантных ДНК в состав генома прокариотов и простейших.
РОЛЬ ОТЕЧЕСТВЕННЫХ УЧЕНЫХ В РАЗВИТИИ МИКРОБИОЛОГИИ
Отечественные исследователи обогатили мировую микробиологию своими выдающимися открытиями. Так, Д. С. Самойлович (1744—1805), испытав на себе, первым применил при-
14
вивки против чумы и разработал методы борьбы с этой грознойинфекцией.
Огромный вклад в развитие отечественной медицины и микробиологии внес современник Л. Пастера и И. И. Мечникова —
Л. С.Ценковский. В 1883 году по аналогии с методом аттенуации Пастера он разработал и получил вакцину против сибирской язвы,которая в течение 60 лет применялась в нашей стране для профилактики заболевания у животных. Л. С. Ценковский по праву считается основателем медицинской микробиологии в России.
В конце XIX века возникла сельскохозяйственная микробиология, основоположником которой был С. Н. Виноградский. В 1890 году он открыл нитрифицирующие бактерии и вместес В.Л. Омелянским доказал их роль в круговороте азота в природе. Основоположником учения о заразных болезнях является отечественный ученый Г. Н. Минх (1836—1896). Он первым обнаружил в крови больных бациллы сибирской язвы и, в противоположность существующему в то время мнению о ее наследственом происхождении, показал инфекционную природу сибирской
Выдающийся ученик И. И. Мечникова Н. Ф. Гамалея первым освоил методику приготовления вакцины против бешенства и применил ее в России, организовал первую пастеровскую станцию в Одессе для прививок населения от бешенства, провел крупные исследования по изучению туберкулеза, бешенства, холеры.
В 1898 году Н. Ф. Гамалея впервые наблюдал явление бактериофагии, провел фундаментальные исследования по изучению невосприимчивости организма и создал одну из теорий иммунитета, обосновал применение так называемых химических вакцин — профилактических препаратов, признанных в настоящее время наиболее перспективными. Значителен вклад Н. Ф. Гамалеи в производство и применение вакцин для профилактики оспы, сыпного тифа, чумы и т. д.
Л. А. Тарасович разработал методы борьбы с сыпным тифом,
доказал эффективность прививок против туберкулеза, внес огромный вклад в изучение механизмов действия ферментов фагоцитов. Российский государственный научно-контрольный институт медицинских биологических препаратов носит имя Л. А. Тарасовича.
Особо отягощенным по эпидемиологическим и инфекционным показателям стал период революции 1917 года, гражданской войны и последующего восстановления народного хозяйства. Активное участие в организации противоэпидемической борьбы принимали Д. К. Заболотный, Л. В. Громашевский, М. Н. Соловьев.
После окончания гражданской войны отечественными микробиологами выполнены фундаментальные исследования, ставшие этапными в истории отечественной и мировой микробиологии.
15
Д. К. Заболотным, Н. Н. Жуковым-Вережниковым, М. П Пок- ровской, С. И. Коробковой, В. М. Туманским были получены эффективные вакцины против чумы, разработана схема лечениячумы. В 30-х годах 3. В. Ермольева и Г. Ф. Гаузс возглавили рабо- ты по изысканию и получению антибиотиков. 3. В. Ермольевойпринадлежит приоритет получения первого кристаллического пе- нициллина
В стране сформировались крупные научные школы с мировым именем. Это, прежде всего, школы вирусологов — Л. А. Зильбера, который разработал вирусогенетическую теорию происхождения опухолей, В. М. Жданова, М. П. Чумакова, А. А. Смородинцева, П. Ф. Здродовского.
Отечественные микробиологи, находясь на передовых рубе- жах борьбы с инфекционными заболеваниями, сформировалипрофилактическое направление медицины, которое получило все- мирное признание и легло в основу международных программВсемирной организации здравоохранения.
ОБЩАЯ МИКРОБИОЛОГИЯ
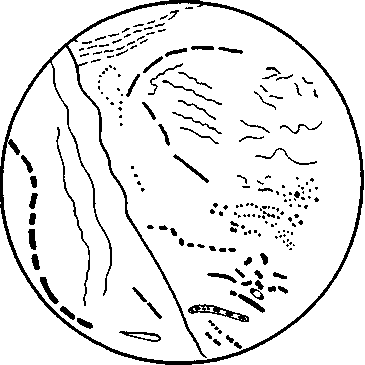
Морфология микроорганизмов
Мир микробов весьма разнообразен и не образует единую систематическую группу. Все микроорганизмы объединены тремя общими признаками:
имеют чрезвычайно малые размеры, колеблющиеся от десятых долей до десятков, иногда сотен микрометров;
большинство микроорганизмов — одноклеточные существа. Встречаются и многоклеточные, но дифференциация клеток у них отсутствует или выражена слабо;
малые размеры микробов определяют специфические, сходные для всех микроорганизмов методы исследования и технику культивирования.
Как известно, основной структурной единицей живого, за исключением вирусов, является клетка; при этом на основании глубоких различий в ультраструктуре различают прокариотический и эукариотический типы организации клеток. Таким образом, все организмы по принципу строения подразделяются на три царства: прокариоты (Рrocariota), эукариоты (Eucariota) и вирусы (Vira).
К прокариотамотносят фотобактерии и скотобактерии, кэу-кариотам —животные, растения, грибы, простейшие. В третье царство выделенынеклеточные формы жизни —вирусы и вироиды.
Все микроорганизмы с прокариотическим типом строения объединены в царство Рrocariota, состоящее из двух отделов: фотобактерии (цианобактерии, или сине-зеленые водоросли) и скотобактерии. К первому отделу относят только сапрофитные формы;
они не являются предметом нашего изучения. Среди скотобактерии, обитающих в организме человека и животных, существуют патогенные виды, составляющие предмет медицинской микробиологии.
Скотобактерии подразделяются на три класса. К классу Васteriaотносятся основные виды бактерий: кокки, палочки, извитые формы и актиномицеты. КлассRickettsiае объединяет облигатных внутриклеточных паразитов — риккетсий и хламидий. Класс
18
Моlliсutes составляют скотобактерии, не имеющие клеточной стенки — микоплазмы.
В соответствии с Международным кодексом номенклатуры бактерий приняты следующие таксономические категории царства прокариот:
ОТДЕЛ - КЛАСС - ПОРЯДОК — СЕМЕЙСТВО - РОД - ВИД.
Для обозначения вида бактерий используется двойная (бинарная) номенклатура, то есть название бактерий состоит из родового и видового обозначений. Вид может подразделяться на подвиды или варианты: биовары(по биологическим свойствам),серовары(по антигенной структуре),фаговары(по чувствительности к фагам).
В практической микробиологии используют более узкие, чем вид, понятия: «штамм» и «клон». Штаммы —это различные микробные популяции одного и того же вида, выделенные из разных источников (организмов людей, животных, объектов окружающей среды) или из одного источника в разное время.Клоном называют популяцию микроорганизмов одного вида, полученную из одной клетки.
ЦАРСТВО ПРОКАРИОТЫ (РКОСАКЮТА)
Ультраструктура прокариотической клетки
Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов —прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
19
геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер — пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток — бактериальные споры;
клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет — до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.
20
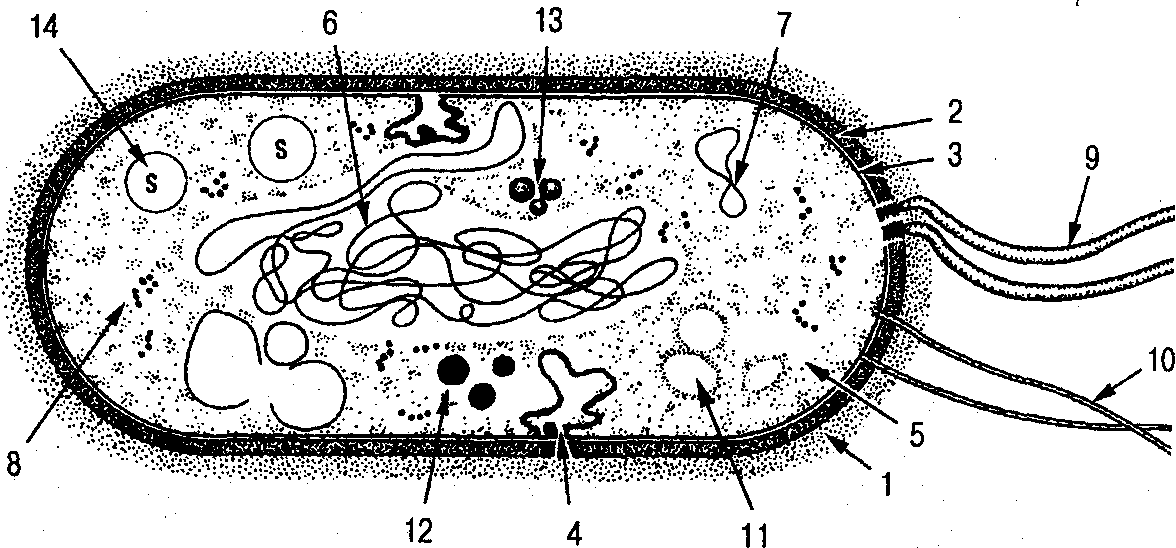
Рис. 1. Схема строения прокариотической клетки:
/ — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — мезосомы; 5 — цитоплазма; 6 — нуклеоид; 7 — плазмида;
8 — рибосомы и полисомы; 9 — жгутики; 10 — пили; 11 — гранулы гликогена; 12 — липидные капельки; 13 — гранулы волютина; 14 — включения серы
Поверхностные структуры.Капсула —это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) —это прямые цилиндрические образования белковой природы длиной 0,3—10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики —органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3—12 мкм и толщиной 10—30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и располо-
21
жение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2—30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка —один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм иL-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01—14 мкм. Она представляет собой плотную эластичную структу -
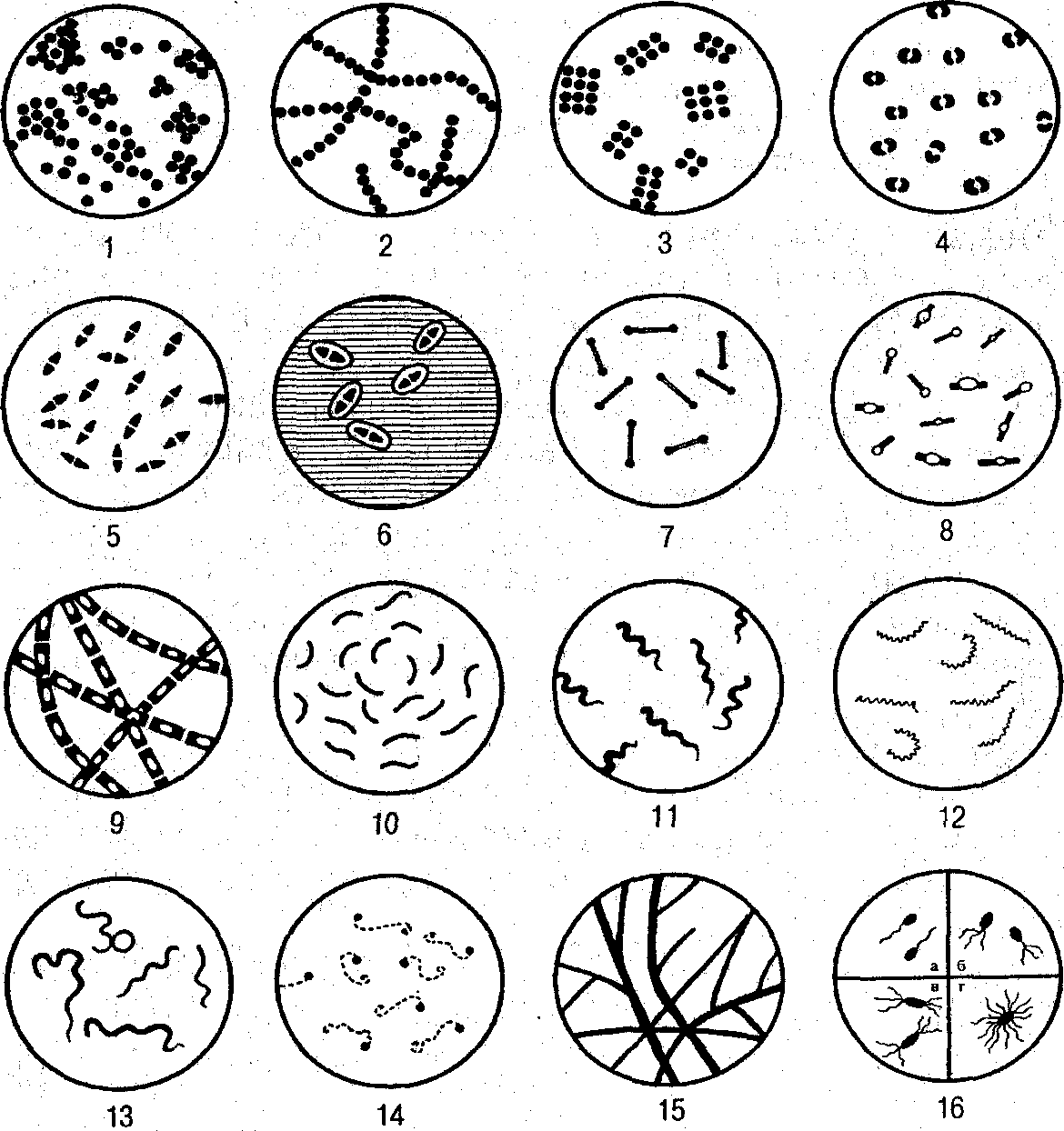
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ — стафилококки; 2 — стрептококки; 3 — сарцины; 4 — гонококки;
5— пневмококки; 6— капсулы пневмококков; 7— коринебактерии дифтерии; 8 — клостридии; 9 — бациллы; 10 — вибрионы; 11 — спириллы; 12 — трепонсмы; — боррелии; 14 — лептоспиры; 15— актиномицеты; 16 — расположение жгутиков: а — монотрихи; б — лофотрихи; в — амфитрихи; г — перитрихи
22
ру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер — пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы.Цитоплазматическая мембрана(ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7—13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15—30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50—70%). В мембране имеются также углеводы (2—5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы —впячивания ЦМ — смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазмазаполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70—80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характер-
23
ными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид —ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2—310в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды —необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы —органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S(так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц — 30Sи 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80Sрибосом.
Споры (эндоспоры) бактерий —покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключен-
24
Иной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора — это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий — бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения.Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других — запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы — гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.
Основные формы бактерий
Основная масса бактерий представлена одноклеточными организмами. Но нередко клетки после деления не расходятся и образуют сочетания различной формы. Эти сочетания неравноценны многоклеточным организмам, поскольку каждая клетка в них автономна и может существовать самостоятельно.
Все бактерии, за исключением микоплазм, имеют определенную форму клетки, поддерживаемую ригидной клеточной стенкой. Для некоторых видов характерен полиморфизм, возникающий под влиянием условий культивирования и факторов внешней среды.
Морфологические типы бактерий немногочисленны. Значительная часть скотобактерий имеют цилиндрическую, сферическую или извитую формы. Основные формы микроорганизмов, относящихся к классу Васteria, приведены на рис. 2.
Кокки,или сферические формы бактерий имеют диаметр 0,5—1 мкм и строго шаровидную, эллипсоидную, иногда коническую формы. В зависимости от взаимного расположения клеток, обусловленного плоскостями деления и сохранением связи между ними, различают несколько морфологических форм.
Одиночно расположенные шаровидные клетки называются микрококками. Кокки, делящиеся в одной плоскости и одном направлении, образуют пары (диплококки) или цепочки (стрептококки). При делении в двух взаимно перпендикулярных плоскостях возникают группы из четырех клеток (тетракокки), а в трех взаимно перпендикулярных плоскостях — пакеты правильной формы из 8—16 клеток (сарцины). При неравномерном делении в нескольких плоскостях наблюдаются скопления неправильной формы, напоминающие гроздь винограда (стафилококки). Патогенные для человека кокки относятся к диплококкам (гонококки, менингококки, пневмококки), стрептококкам и стафилококкам.
Палочки,или палочковидные бактерии имеют цилиндрическую форму, длина их составляет 1—8, а толщина — 0,5—2 мкм; это самая многочисленная группа бактерий, отличающаяся большим морфологическим разнообразием. Их различают по форме конца клетки, который может быть закругленным, утолщенным, обрубленным; взаимному расположению в мазке — беспорядочному, попарному (диплобактерии, диплобациллы), в виде цепочки (стрептобактерии, стрептобациллы); спорообразованию — неспорообразующие палочковидные формы (бактерии) и спорообразующие (бациллы, клостридии).
Извитые бактериибывают трех типов: вибрионы, спириллы и спирохеты.
Вибрионы — слегка изогнутые палочки, похожие на запятую (например, холерный вибрион).
26
Спириллы имеют форму нити с несколькими правильными завитками, среди них известен один патогенный вид, вызывающий болезнь содоку.
Спирохеты — очень подвижные тонкие и длинные клетки, имеющие вид спирали. При отсутствии жгутикового аппарата характерными для них являются различные типы движения: волнообразные, поступательные, винтообразные, маятникообразные. Особенностями ультраструктуры клетки спирохеты является наличие трех структурных элементов: наружной оболочки, соответствующей клеточной стенке, осевой нити (аксостиля), со-стоящей из микрофибрилл, и цитоплазматического цилиндра, винтообразно закрученного вокруг осевой нити. Фибриллы равномерно располагаются по краям клетки, прикрепляясь одним концом к цитоплазматическому цилиндру; в середине клетки они перекрывают друг друга свободными концами. Очевидно, их сокрашение или вращение обусловливает движение спирохет.
Спирохеты, играющие роль в инфекционной патологии человека, относятся к трем родам — Тгероnema,Leptospira, Воггеliа.
L-формы микроорганизмов и их роль в патологии человека
Способность к L-трансформации — свойство, присущее многим видам бактерий.L-формы образуются независимо от их видовой принадлежности при воздействиях, блокирующих синтез основных компонентов клеточной стенки, либо при ее разрушении соответствующими ферментами в условиях повышенной осмотической концентрации среды.
Известен L-трансформирующий эффект пенициллина. Механизм действия этого антибиотика связан с нарушением перекрестного соединения пептидных цепей муреина, обеспечивающего ригидность клеточной стенки бактерий. Муреин состоит из повторяющихся мукопептидных единиц и единиц пептидогликана.
L-варианты могут индуцироваться пенициллином у грамположительных и грамотрицательных бактерий.
В результате изучения способности некоторых антибиотиков индуцировать образование L-форм бактерий антибиотики условно разделяют на три группы:
универсальные индукторы L-форм, вызывающие их образование независимо от видовой принадлежности микроорганизмов (пенициллин);
антибиотики, оказывающие избирательное L-трансформирующее действие в зависимости от видовой принадлежности микроорганизма. Например, бацитрацин вызывает индукциюL-форм стрептококков; бацитрацин, ванкомицин и ристомицин индуцируют образованиеL-форм N.meningitidis. Стрептомицин являет-
27
ся активным фактором индукции L-форм молодых культур холерного вибриона иMycobacterium tuberculosis;
к антибиотикам, действие которых не вызывает L-трансформирующего эффекта, относятся тетрациклин, канамицин, хлорамфеникол и др.L-формы некоторых грамположительных бактерий нередко образуются при комбинированном воздействии лизоцима, пенициллина и глицина.
При изучении действия разнообразных химических препаратов установлено, что соли ртути, кадмия, хрома, лития, а также фенол и формальдегид практически не вызывают индукции L-форм. Из физических факторов, вызывающихL-трансформацию, можно отметить действие ультрафиолетовых лучей.
Непременным условием индукции L-вариантов бактерий является включение в состав среды нормальной сыворотки млекопитающих, которая обладает инактивирующим действием на те вещества питательных сред, которые ингибируют ростL-форм.
Морфология L-вариантов бактерий изучается с помощью световой микроскопии, окрашенных препаратов, фазово-контрастной микроскопии с серийной и цейтрафферной киносъемкой, люминесцентной микроскопии с негативным контрастированием и ультратонкими срезами.
Нестабильные L-формы бактерий имеют две периферические мембраны, из них наружная, очевидно, представляет деградированную клеточную стенку, а внутренняя — цитоплазматическую мембрану. СтабильныеL-формы имеют только цитоплазматическую мембрану.
Цитоплазма L-форм структурно сходна с цитоплазмой интактных бактерий, но уL-форм в ней имеются большие вакуоли и гранулы внутри вакуолей. УL-форм мезосомы утрачиваются и происходит непосредственное прикрепление нуклеоида к мембране. Вследствие этогоL-формы теряют клеточную стенку, иногда сохраняя измененные ее фрагменты; отмечается причудливость конфигурации мембран и наличие множества телец и волокон, содержащихся в пузырьках, ограниченных мембраной. Структурные элементыL-форм подразделяют на простые и комплексные. Их размеры варьируют от крупных (10 мк) до субмикроскопических гранул, фильтрующихся форм (250 ммк). Способность L-форм прорастать через мелкие поры бактериальных фильтров связана не только с их размерами, но и с пластичностью — крупные структуры, легко деформируясь, проходят через поры более мелких фильтров.
Микроструктуры L-форм представлены РНК- и ДНК-содержащими элементами, преобладают последние.
L-формы иногда сохраняют некоторые виды ферментативной активности. Например, некоторые штаммы L-форм стрептококка продуцируют 0-стрептомицин, стрептокиназу, ДНК-азу,
28
М-белок; L-формы холерных вибрионов продуцируют нейраминидазу, L-формы Сl.tetani— столбнячный экзотоксин.
В связи с отсутствием клеточной стенки L-формы имеют антигенные особенности. У L-форм преобладают антигенные детерминанты цитоплазматической мембраны и цитоплазмы.
Способность бактерий культивироваться в L-форме независимо от наличия в среде L-трансформирующих агентов называется стабилизацией. При этом происходит необратимая утрата определенных звеньев биосинтеза клеточной стенки и способности восстановления. Нестабильные L-формы отличаются тем, что при их культивировании на средах, не содержащих индуцирующего фактора, происходит реверсия бактерий исходного вида.
Вопрос о природе наследственных механизмов, обусловливавших индукцию, стабилизацию и реверсию L-форм бактерий, мало изучен. Вероятно, превращение в L-формы и их реверсия могут происходить в результате мутаций. Кроме мутационного механизма существует массовая конверсия L-форм в результате непосредственного воздействия разных агентов на клеточную стенку.
В 1944 году Dienes иSmith выделили L-формы бактерий из организма больного перитонитом. С 1953 года появились сведения о выделении L-форм стрептококка из крови больных септическим эндокардитом, гнойным менингитом и менингоэнцефалитом, а также в периоды ремиссии при пиелонефрите.
Образование L-форм и близких вариантов бактерий под влиянием антибиотиков и других факторов в организме, длительная их персистенция и возможность реверсии исходных видов бактерий доказывает, что L-формы далеко не безразличны для организма хозяина. Имеются данные о L-формах, сохранивших исходную степень патогенности, например, вирулентные штаммы L-форм холерного вибриона, Сlostridium tetani, С1.perfringens. Способность L-форм продуцировать ферменты агрессии, экзо- и эндотоксиныcвидетельствует о сохранении многих факторов вирулентности.
Видовая идентификация L-форм посредством определения вида бактерий, реверсировавших из L-форм, затруднена в связи с неполным восстановлением при этом признаков исходного вида. Все это усложняет микробиологическую диагностику, течение инфекционного процесса, ведет к рецидивам инфекции, бактерионосительству, снижает эффективность лечения и создает очаги инфекции, не поддающиеся эпидемиологическому контролю.
Морфология и биологические свойства
микоплазм
Микоплазмы — наиболее мелкие полиморфные микроорганизмы, живущие в условиях искусственных питательных сред и не имеющие клеточной стенки. Из-за отсутствия клеточ-
29
ной стенки микоплазмы полиморфны, их размеры — 50—300 нм, форма — в виде шаров, колец, нитевидных форм, зерен.
Большинство микоплазм неподвижно, однако у некоторых (Мусорlasma рneumoniaе) обнаружены структуры, которым приписывают функцию движения. Микоплазмы не образуют спор и капсул, слабо окрашиваются анилиновыми красителями, для окраски используется метод Романовского — Гимза.
Микоплазмы выделены в класс Моllicutes («мягкокожие»). Единственным критерием объединения этих микроорганизмов в класс является отсутствие клеточной стенки и ее предшественников.
По вопросу филогенетического происхождения микоплазм существуют несколько точек зрения. М. А. Пешков (1969) считал, что именно микоплазмы явились предками прокариотов, в том числе и бактерий, а способность к синтезу клеточной стенки является следствием серии мутаций. Другая гипотеза Еdvard иFreundt (1969) предполагает отсутствие родства между микоплазмами и бактериями на основании наличия у всех стеринзависимых микоплазм холестерина, включенного в мембрану, отсутствия генетического родства микоплазм и их возможных бактериальных предков в опытах по молекулярной гибридизации ДНК-ДНК и ДНК—РНК, а также различные, по мнению авторов гипотезы, способы репродукции микоплазм иL-форм бактерий.
В класс Моllicutesвходит один порядок Мусорlasmatales, в составе которого известны два семейства. Стеринзавиcимые микоплазмы объединены в семейство Мусорlasmataсеае с одним родом Мусорlasma, куда входят 33 вида микоплазм человека и животных. Стериннезависимые микоплазмы объединены в семейство Асhо-leрlasmataсеае, состоящее из одного рода семейство Асhоleрlasma, и в настоящее время в него входят три вида.
На плотных питательных средах микоплазмы растут в виде характерных колоний с уплотненным, врастающим в среду центром и нежным краем, по форме напоминают яичницу-глазунью — «friend egg». В условиях жидких и полужидких сред рост происходит в виде нежного диффузного помутнения.
В зависимости от характера питательных сред микоплазмы имеют такие морфологические особенности: при выращивании на плотных питательных средах они представлены пластическими сгустками протоплазмы неопределенной формы, а в жидких средах — разнообразием форм — кольцами, нитями, гранулами, палочковидными и спиральными образованиями.
Одним из наиболее значимых факторов стабильности и эластичности мембраны микоплазм является холестерин — главный липидный компонент у так называемых паразитических микоплазм. Содержание значительного количества холестери-
30
на сближает мембраны микоплазм с мембранами животных клеток, хотя по ферментативной активности мембраны микоплазм ближе к мембранам бактериальных протопластов.
Патогенные микоплазмы могут вырабатывать экзотоксин. Например, М.neurolyticumвыделяет истинный экзотоксин, который является термолабильным белком и быстро прикрепляется к рецепторам астроцитов мозга. По способности продуцировать |вемолизин микоплазмы подразделяются на виды, вызывающие β-гемолиз, например М.laidlauii, М.pneumoniae, α-гемолиз, и не вызывающие гемолиза.
Вид М.mycoides var. mycoides отличается от других микоплазм наличием галактановой капсулы; экстрагированный галактан по твоему воздействию сходен с эндотоксином грамотрицательных бактерий. Микоплазмы имеют ряд ферментов, нарушающих нормаль-метаболизм клеток. Так, аргининдегидролазные ферменты (разрушают необходимый для жизни клеток аргинин. Нуклеозид-фосфорилаза — фермент микоплазм, который вызывает расщепление в клетках тимидина, что нарушает их нормальное размножение.
Ультраструктурные изменения в клетках, индуцированные микоплазмами, как при латентной, так и при острой микоплазменной инфекции сходны с соответствующими изменениями, вызываемыми вирусами. Микоплазмы, как и вирусы, вызывают феномен бляшкообразования.
При электронной микроскопии микоплазмы, расположенные внутри клеток, трудно отличить от вирусных частиц. Микоплазмы можно обнаружить и на мембранах клеток хозяина, причем связь такова, что невозможно дифференцировать мембрану клетки от мембраны микоплазм. В цитоплазме клеток при микоплазмоинфекции нередко обнаруживаются гранулярные включения, явления пикноза и кариорексиса.
Многие виды микоплазм являются патогенными для человека, вызывая поражения дыхательных и мочеполовых путей, суставов, сердца, нервной системы. Среди них наибольшее значение в инфекционной патологии человека имеют: М. pneumoniae (возбудитель небактериальной пневмонии, бронхитов» воспаления среднего уха); М.hominis 1 (возбудитель воспалительных процессов верхних дыхательных путей, фарингитов, ангин, шейной аденопатии); М.hominis 1 и М.hominis2 (возбудители уретритов небактериального происхождения); микоплазмы Т-групп|ы (возбудители негонорейных уретритов и других заболеваний мочеполовой сферы).
31
Морфология и биологические свойства риккетсий и хламидий
Риккетсии и хламидий составляют класс Rickettsiaе.Риккетсииотличаются полиморфизмом, их размеры варьируют от 0,4 до 10—40 мкм. Выделяют кокковидную, палочковидную, нитевидную формы. Они не имеют жгутиков, не образуют спор и капсул, окрашиваются по Граму, Романовскому-Гимза, Здродовскому. Эта группа прокариотов является облигатными внутриклеточными паразитами различных групп животных и человека. У человека риккетсий вызывают сыпной тиф,Q-лихорадку, пятнистую лихорадку скалистых гор и другие риккетсиозы.
Хламидии— облигатные внутриклеточные организмы прока-риотной природы. Изучение ультраструктуры хламидий с помощью электронного микроскопа началось с конца 40-х годов XX века.
Цикл развития хламидий представлен двумя основными формами, сменяющими друг друга — ретикулярные тельца (вегетативные формы) и элементарные тельца (спороподобные формы).
К виду Сl.trachomatis относят патогенные для человека возбудители трахомы, урогенитальных хламидиозов, венерической лимфогранулемы, мышиной пневмонии.
На основании уникальности цикла развития хламидий они были выделены в особый порядок Сlamydiales, семейство Сlamydiasсеае, род Сlamydia, объединяющий два вида: Сl.trachomatis, Сl.psittaci.
Хламидии являются возбудителями трахомы, венерического лимфогранулематоза. К виду Сl.psittaci отнесены возбудители пситтакоза (орнитоза), пневмонии, полиартритов, энтеритов, конъюнктивитов, энцефалитов домашних, сельскохозяйственных и диких животных. Нередко эти микроорганизмы вызывают хламидиозы у людей с возможным распространением инфекции от человека к человеку.
Основа цикла развития хламидий — закономерная смена вегетативных репродуцирующих неинфекционных клеток (ретикулярных телец — РТ) спороподобными инфекционными клетками (элементарными тельцами — ЭТ). Эти две формы различаются по ультраструктуре, но сходны у разных штаммов хламидий.
Морфологически вегетативные формы хламидий сходны с некоторыми грамотрицательными бактериями, например, возбудителем туляремии. Они округлой формы, диаметром до 1—1,5 мкм, окружены клеточной стенкой и цитоплазматической мембраной, имеющей трехслойную структуру.
Спороподобные клетки хламидий не имеют морфологических аналогов среди бактерий и представляют собой мелкие, диаметром 0,25—0,3 мкм, сферические клетки с плотным эксцентричным нуклеоидом.
32
Вегетативные ретикулярные клетки обладают выраженной метаболической активностью. Спороподобные хламидий отличаются весьма слабым метаболизмом.
Прохождение цикла развития является обязательным условием внутриклеточного паразитирования хламидий, без него невозможно образование нового поколения возбудителя.
Зрелой морфологической структурой являются элементарные тельца - они грамотрицательны и обладают инфекциозностью.
Основные этапы цикла развития микроорганизма: контакт ЭТ с поверхностью чувствительной клетки; проникновение ЭТ в цитоплазму путем фагоцитоза; преобразование ЭТ в вегетативную форму с хрупкой клеточной стенкой; деление РТ, с последовательным уменьшением их размера, реорганизацией внутреннего содержимого и образованием «переходных форм»; их преобразование в ЭТ нового поколения микроорганизма; выход микроорганизма из клетки.
Цикл развития (гальпровий) хламидий происходит в цито-плазматических включениях, определяемых как тельца Гальберштед-тера-Провачека, которые представляют собой ограниченную фагоцитарную вакуоль, содержащую различные морфологические структуры микроорганизма.
Сходство химического состава клеточной стенки хламидий и грамотрицательных бактерий позволяет использовать для выявления телец Гальберштедтера-Провачека световую микроскопию при окраске препаратов по Романовскому-Гимза, а также фазовый контраст при просмотре неокрашенных препаратов.
Хламидии содержат два типа нуклеиновой кислоты (ДНК и РНК). Отличаясь дефицитом ферментных систем, хламидий используют хозяина как поставщика метаболической энергии. Облигатный характер внутриклеточного паразитизма характеризуется как энергозависимый паразитизм. Хламидий не производят собственную АТФ, подавляют синтез ДНК клетки-хозяина.
Резистентноеть хламидий во внешней среде высокая и для заживания водных резервуаров общественного пользования, инфицированных обитающими в урогениталиях человека возбуди-телей паратрахомы, необходимо гиперхлорирование воды. Многие штаммы этих микроорганизмов проявляют высокую чувствительность к сульфаниламидам и антибиотикам широкого спектра действия: тетрациклинам, макролидам, рифампицину. Культивируются в куриных эмбрионах и культурах клеток.
Хламидии содержат стабильный (100˚С) липопротеиновый антиген, расположенный в клеточной стенке. Антигены хламидий обладают слабой иммуногенной активностью. Уровень гуморального иммунитета на инфицирование хламидиями обычно низкий.
33
Морфология и биологические свойства актиномицетов
Актиномицеты относятся к классу Bacteria. Морфоло- гической особенностью актиномицетов является образование од- ноклеточного мицелия — скопления нитевидных выростов (гиф) одной клетки. Свое название актиномицеты получили от первого из описанных видов — Actinomyces bovis — «лучистого грибка», представляющего собой в пораженных тканях радиально распо- ложенные сплетения гиф с колбовидными вздутиями на концах (друзы лучистой формы).
У одних видов актиномицетов (Nocardia, Actinomyces) деле- ние происходит путем распада мицелия на палочковидные и сфе- рические формы, другие (Streptomyces, Actinomyces) размножа- ются, в отличие от бактерий, экзо- и эндоспорами.
Окрашиваются актиномицеты по Граму, являясь грамполо- жительными, что свидетельствует об общности химического со- става их клеточной стенки с другими прокариотами.
Большинство видов являются сапрофитными, многие из них — продуценты антибиотиков. Патогенные виды вызывают у челове- ка актиномикоз и нокардиоз.
ЦАРСТВО ЭУКАРИОТЫ (EUCARIOTA)
}
Морфология грибов
Грибы — большая группа эукариотических организ- мов, объединяющая свыше 100 000 видов. В систематике органи- ческого мира грибы занимают особое положение. С животными грибы сближает наличие в оболочке их клетки полисахаридной субстанции — хитина (исключение составляют оомицеты, у кото- рых обнаружена целлюлоза), участие в обмене азота — мочевины, а в обмене углеводов — гликогена. Только в клетках животных и грибов имеются цитохромы, участвующие в окислительно-вос- становительных процессах. Однако по способу питания (адсорб- тивное — путем всасывания, а не заглатывания пищи) и неогра- ниченному росту грибы напоминают растения. Они не содержат хлорофилла и по типу питания являются гетеротрофами: сапро- фитические грибы используют остатки растительного или живот- ного происхождения, а паразиты — ткани растений и животных Проблемы систематики грибов до сих пор нельзя считать раз- решенными. Рассматривается вопрос о выделении грибов в са- мостоятельное царство, но в настоящее время пользуются бота- нической классификацией. В ее основу положен морфологический принцип строения тела грибов и органов полового воспроизве- дения в культуре. Согласно этому грибы разделяются на выс-
34
щие и низшие, совершенные и несовершенные. Различают пять классов:
архимицеты (Archimycetes);
фикомицеты (Phykomycetes) с тремя группами: хитридиомицеты (Chitridiomycetes), оомицеты (Oomycetes), зигомицеты (Zygomycetes);
аскомицеты (Ascomycetes);
базидиомицеты (Basidiomycetes);
дейтеромицеты (Deuteromycetes).
Архимицеты и фикомицеты относятся к низшим грибам. Пер- вые из них мицелия не имеют, мицелий вторых несептирован. Эти грибы обладают наиболее простой формой спороношения в виде спорангиев, в которых заключены споры.
У аскомицетов и базидиомицетов, являющихся высшими гри- бами, мицелий септированный (многоклеточный), строение ор- ганов спороношения более сложное.
Представители первых четырех классов имеют половой и бес- полый циклы развития и называются совершенными.
Дейтеромицеты являются высшими несовершенными гриба- ми: они обладают септированным мицелием, но половое споро- ношение у них отсутствует или не обнаружено. Группа несовер- шенных грибов непостоянна, поскольку обнаружение у гриба половых форм размножения позволяет отнести его к одному из перечисленных классов.
Патогенные для человека грибы — возбудители микозов (от греч. mykes — гриб) относят к различным систематическим груп- пам ботанической классификации.
Грибы различают по величине, строению, местам произрас- тания и физиологическим функциям. Их размеры варьируют от микрометров (микроскопические грибы) до метров (шляпочные). Исходя из особенностей питания и мест произрастания сформи- рованы различные экологические группы: почвенные, фитопато- генные, энтомофилы, зоофилы, антропофилы и др.
Наряду с различиями грибы имеют общие черты строения. Грибы — это своеобразные эукариотические организмы, которые в процессе своего развития претерпевают морфологические изме- нения. Данные изменения могут быть связаны с жизненнымициклами — половым и бесполым или с влиянием факторов внеш- ней среды. Основу клеточного строения большинства из них со- ставляет масса тонких, ветвящихся трубчатых нитей, именуемых гифами, а вся эта масса гиф называется мицелием (рис. 3). Диа- метр гиф варьируется от1 до10 мкм, а их длина— от4-6 до80—100 мкм и более. Каждая гифа окружена тонкой жесткой стен- кой, основным компонентом которой является хитин — азотсо- держащий полисахарид, являющийся, как известно, структурным
![]()

1 — несептированный (одноклеточный) мицелий; 2— септированный (многоклеточный) мицелий; 3 — псевдомицели и
компонентом наружного скелета членистоногих. Гифы не имеют клеточного строения. Протоплазма гиф либо совсем не разделена (у одноклеточных), либо разделяется поперечными перегородками, называемыми септами (у многоклеточных). Такие септы делят содержимое гиф на отдельные участки, внешне похожие на клетки, при этом образование септ не связано с делением ядер. В центре септы, как правило, остается небольшое отверстие (пора), через которое протоплазма может перетекать из одного компартмента в другой. В каждом компартменте могут находиться одно, два или несколько ядер. Гифы, не имеющие перегородок, образуют несептированный мицелий, а имеющие — септированный.
У дрожжевых и дрожжеподобных грибов образуется псевдомицелий, который в отличие от истинного мицелия, представляющего собой ветвистую трубку, разделенную у высших грибов поперечными перегородками, не имеет общей стенки, является цепочкой из клеток, которая формируется в процессе размножения почкованием.
Основой клеточной стенкигрибов в отличие от бактерий являются полисахариды, состоящие из пяти-шести моносахаров. В связи с их меньшей, чем у бактериальных оболочечных белков, вариабельностью, наблюдается антигенное родство между далекими по своим морфологическим свойствам видами грибов. В состав полисахаридных фракций входят глюкозамин, манноза, глюкоза, ксилоза.
Непосредственно к внутренней части клеточной стенки прилегает цитоплазматическая мембрана,с которой в тесном контакте находитсяцитоплазматический ретикулюм,часто гранулярный, составляющий основную частьцитоплазмы.В ней расположены одно или несколькоядер,имеющих свою оболочку с порами, иядрышко, содержащее в составе хромосом ДНК. В цитоплазме естьцентральная вакуоль,а такжемитохондрии, микросомы, лизосомы, рибосомы, пластиды, комплекс Голъджи, глико-, липо-, хромопротеиды, секреторные гранулы, миелоидные образования,различные включения[волютин, гликоген, пигментыи др.).
Кроме того, в клетках могут накапливаться продукты метаболизма грибов — антибиотики, ферменты, витамины, токсины,и т. п.
36
При росте на питательной среде мицелий ветвится боковыми выростами гиф; переплетаясь, он образует грибницу. Различают мицелий вегетативный — погруженную в субстрат питающую часть грибницы; репродуктивный — воздушную, спороносящую часть грибницы; склероций — покоящийся тип грибов. Последний представляет собой округлое или продолговатое тело плотной консистенции диаметром от нескольких миллиметров до десятков сантиметров, образующееся веществами тесного переплетения гиф. Он богат запасными питательными веществами и обеспечивает жизнеспособность гриба в неблагоприятных условиях внешней среды (недостатке влаги, низких температурах и т. п.).
Различают два типа размножения грибов — половоеибесполое.По способу осуществления размножение может бытьвегетативным,то есть происходить без образования специальных или или с помощью малодифференцированных органов размножения;репродуктивньш —путем образования специальных органов воспроизводства. Во втором случае возможно как бесполое, так и половое воспроизводство.
Вегетативное размножение грибов может осуществляться ку-сочками мицелия, склероциями, спорами, возникающими в результате расчленения гиф мицелия (хламидоспоры, артроспоры, бластоспоры, оидии, геммы).
Репродуктивное размножение грибов происходит при посредстве спор, возникающих половым или бесполым путем на специальных дифференцированных или спороносящих ветвях мицелия или органах.
Бесполое размножение осуществляется при помощи спор, развивающихся эндогенно (эндоспоры, спорангиеспоры) и экзогенно — на концах особых выростов мицелия или на мицелии (конидии).
Половое размножение у грибов осуществляется путем слияния мужских и женских половых гамет, в результате чего образуется зигота. Способы полового размножения у грибов весьма разнобразны. У низших грибов происходит слияние одинаковых (изогамия) или разных по размерам (гетерогамия) гамет. Из зиготы после периода покоя вырастает спорангиеносец со спорангием, наполненным эндоспорами. У высших грибов образуются для полового размножения аскоспоры и базидиоспоры.
У несовершенных грибов половой процесс заменяется гете-рокариозом (разноядерностью) и парасексуальным процессом |без слияния ядер в первом случае или со слиянием ядер во втором).
Патогенные для человека микроскопические грибы относятся к зигомицетам, аскомицетам, дейтеромицетам.
Зигомицеты —группа грибов класса фикомицетов; это назем-ные, чаще всего почвенные сапрофиты, мицелий в основном не-
37
септированный. Имеют особый тип полового процесса — слияние двух недифференцированных на гаметы клеток, с образованием покоящейся зигоспоры, с диплоидным набором хромосом. Бесполое размножение зигомицетов происходит с помощью эндоспор, образующихся в специальных крупных шаровидных клетках — спорангиях, которые формируются на свободных концах плодоносящих гиф — спорангиеносцах, имеющих разнообразную форму: грушевидную ( Mucor), шаровидную (Rhizopus), булавовидную (Асtinomucor) и др. Каждая спора дает начало новому мицелию.
Грибы родов Mucor,Rhizopus, Аbsidia, среди которых наиболее широко известен мукор, могут вызывать микозы у человека и животных, связанные с поражением кожи и легких.
Класс аскомицетов —одна из самых многочисленных и сравнительно высокоорганизованных групп микроскопических грибов, отличающаяся от зигомицетов сложностью строения, особенно репродуктивных органов. К ним относятся дрожжи и многие виды плесневых грибов, последние имеют расчлененный многоклеточный мицелий.
У аскомицетов в результате полового процесса, которому предшествует плазмогамия (слияние половых клеток), кариогамия (слияние ядер половых клеток) и мейоз, формируются специализированные клетки округлой формы — аски (сумки). В сумке образуется определенное число (чаще всего восемь) аскоспор, с помощью которых и происходит размножение. У одних аскомицетов сумки возникают непосредственно на мицелии, у других — внутри или на поверхности плодовых тел, образуемых в результате сплетения гиф мицелия.
Аспергиллы и пенициллы образуют сумки чаще всего округлой формы. У большинства аскомицетов плазмогамия и кариогамия при половом процессе разделены во времени. Бесполое размножение у дрожжей осуществляется почкованием, а у мицелиальных форм грибов — с помощью экзоспор (конидий), образующихся на конидиеносцах. Грибы рода Аspergillus и рода Реnicillium отличаются строением органов вегетативного спороношения. У аспергиллов на конидиеносцах находятся удлиненные клетки (стеригмы), которые формируют цепочки экзоспор — конидий. Плодоносящая гифа пенициллиума имеет три ряда стеригм, на которых располагаются округлые конидии в виде цепочек.
Среди аспергиллов и пенициллов встречаются антибиотико-образующие виды и патогенные для человека. Последние являются возбудителями таких микозов как аспергиллез и пенициллиоз (поражение кожи, слизистых, легких).
Дейтеромицеты —несовершенные грибы (Fungi imperfecti), объединяют более 25 000 видов, с септированным мицелием, весь их жизненный цикл проходит в гаплоидной стадии, без смены ядерных фаз. Мицелиальные грибы размножаются только с по-
38
мощью конидий. Из патогенных для человека к ним относятся возбудители дерматомикозов: трихофитии, микроспории, эпидер-мофитии, фавуса. Поражают кожу и ее придатки (волосы, ногти), вызывают тяжелые, склонные к хроническому протеканию заболевания, сопровождающиеся аллергией.
Морфологически дерматофиты очень разнообразны в зави-симости от условий обитания. В таких тканях как кожа и ногти дерматофиты представлены нитями мицелия, а в волосах — спорами с различным типом расположения (снаружи и внутри волоса). На питательной среде они имеют септированный мицелий с его различными видоизменениями (спиралями, «канделябрами», «оленьими рогами» и т. п.) и разные виды конидий — макро- и микроконидии.
К несовершенным грибам относятся и дрожжеподобные грибы рода Саndidа, образующие в результате почкования псевдомицелия; у патогенных видов Саndidа присутствуют также покоящиеся формы в виде хламидоспор (толстостенных округлых, толненных липидами клеток, образовавшихся из вегетативных). Эти грибы являются условно-патогенными и могут вызывать поражение кожи, слизистых оболочек и внутренних органов (кандидозы или кандидомикозы). Поскольку грибы рода Саndidа являются представителями нормальной микрофлоры человека, поражение происходит чаще всего эндогенным путем на фоне снижения иммунной реактивности, дисбактериозов, нарушения обмена веществ, длительном применении антибактериальных средств широкого спектра действия.
Морфология простейших
Простейшие (Рrotozoa) представлены одноклеточными эукариотическими организмами, относящимися к животным. Они
широко распространены в природе (около 2500 видов) и ведут
свободный или паразитический образ жизни.
Простейшие играют значительную роль в инфекционной па-тологии человека и животных. Некоторые из них являются без-вредными обитателями кишечника (например, кишечная амеба), другие реализуют свою патогенность обычно при массивном за-ражении на фоне иммунодефицитных состояний (лямблии, пнев-моцисты) и, наконец, часть видов представлена тканевыми и кро-вяными паразитами, вызывающими острые или хронические заболевания (лейшмании, трипаносомы, малярийные плазмодии). | Являясь по ультраструктуре эукариотами, простейшие имеют микроскопические размеры (3-150 мкм) и сложное строение: одно или несколько дифференцированных ядер (с ядерной мембраной и ядрышками) и цитоплазму с эндоплазматическим ретикулюмом, митохондриями, аппаратом Гольджи, лизосомами, рибосомами,
39
включениями, пищеварительной и сократительной вакуолями. Ядро содержит набор хромосом, которые после репликации разделяются путем митоза.
В цитоплазме выделяют эндоплазму с органеллами и более уплотненную наружную часть — эктоплазму. У некоторых простейших, например, у саркодовых, лишенная жесткой оболочки протоплазма, переливаясь, образует выросты (псевдоподии или ложноножки), благодаря которым клетки передвигаются и не имеют постоянной формы. Но у большинства простейших периферический слой цитоплазмы имеет эластичную плотную мембрану — пелликулу, поддерживающую определенную форму этих микроорганизмов. Некоторые виды простейших поверх пелликулы формируют жесткую структуру, нередко пропитанную солями кальция, стронция и кремнекислыми соединениями — кутикулу, которая служит дополнительным средством защиты от механических, химических воздействий и высыхания.
Ряд простейших в определенных условиях может превращаться в цисты. Они окружены плотной двухконтурной оболочкой, сходной с кутикулой, и содержат по нескольку ядер. Цисты простейших являются покоящимися формами.
Кроме этого, некоторые простейшие имеют опорные фибриллы (осевые нити, или «аксостиль»), выполняющие функции своеобразного «скелета».
Многие простейшие подвижны. Одни виды передвигаются с помощью псевдоподий, другие снабжены специальными двигательными органеллами — жгутиками и ресничками. Строение жгутиков и ресничек типично для эукариотических структур: на периферии жгутика располагаются девять двойных белковых нитей, а в центре — две одиночные нити (структура 9+2). Снаружи эта система покрыта плазматической мембраной. Основание жгутика закреплено в наружном слое цитоплазмы при помощи базального тельца (блефаропласта). Для окраски простейших чаще всего пользуются методом Романовского — Гимза, при котором цитоплазма клетки окрашивается в голубой, а ядро, блефаропласт и жгутики — в красный цвет.
Простейшие могут размножаться бесполым и половым путем. Размножение некоторых видов бывает весьма сложным, со сменой бесполого и полового циклов.
Бесполое размножение осуществляется по типу простого деления, когда делится ядро, затем — протоплазма и образуются два дочерних индивида; множественного деления (дробления) сначала ядра, а потом и всей клетки на ряд молодых особей (этот вид деления называется шизогонией).
Половой процесс у простейших происходит в форме копуляции или конъюгации, а также путем самооплодотворения — аутогамии, при которой сливаются отдельные ядра.
40
Деление простейших на классы основывается на способах передвижения и особенностях размножения. Тип Рrotozoa составляет четыре класса: жгутиковые (Flagellata); саркодовые (Sarcodina); споровики (Sроrozoa); реснитчатые (Сiliata).
В эволюционном плане примитивными простейшими являются жгутиковые формы. Амебовидные и реснитчатые формы образовались, как считают специалисты, в результате развития предковых жгутиковых. Остановимся на более подробной характеристике патогенных простейших перечисленных классов.
Класс жгутиковые —простейшие с одним или несколькими жгутиками, а в некоторых случаях с ундулирующей (волнообразной, от лат.undula— волна) мембраной, имеющей общую со жгутиками природу, размножаются путем простого продольного деления. В. эту группу входят простейшие, обитающие в кишечнике и мочеполовой сфере (лямблии, трихомонады), а также жгутиковые, паразитирующие в крови или тканях (трипаносомы, лейшмании).
Из лямблийспецифическим паразитом человека являетсяLamblia intestinalis, вызывающая лямблиоз (поражение тонкого кишечника, желчного пузыря и желчевыводящих путей). Клетка лямблии — грушевидной формы с двусторонней симметрией и длиной 10—18 мкм. В более широкой передней части простейшего находится вогнутый (присасывательный) диск, с помощью которого лямблия плотно прикрепляется к эпителию кишечника. В клетке имеются два симметрично расположенные ядра, четыре пары жгутиков и две осевые фибриллы посередине. В отличие от других простейших у лямблий отсутствуют типичные митохондрии, аппарат Гольджи и сократительные вакуоли. В толстом кишечнике вегетативные клетки переходят в стадию цист, которые представляют собой двух-трехъядерные эллиптические формы с толстой оболочкой, длиной 8—12 мкм и шириной 3—10 мкм.
Род трихомонадывключает три паразитических для человека вида: Тrichomonas hominis— обитает в кишечном тракте; Т.tenax— паразиты ротовой полости; Т.vaginalis— поражает урогенитальные пути.
Из перечисленных безусловно патогенным является последний вид. Инфекция, вызываемая трихомонадами, носит название трихомоноз. Эти простейшие имеют грушевидную форму длиной около 15—30 мкм, с четырьмя расположенными пучком спереди жгутиками, один из которых ограничивает внешний край короткой ундулирующей мембраны, и опорной эластичной нитью, проходящей через всю цитоплазму. Стадия цисты у них отсутствует.
Среди патогенных трипаносомразличают возбудителей аф-риканского трипаносомоза, или сонной болезни (Тгураnosoma brusei), передаваемого кровососущей мухой цеце; американско-
41
го трипаносомоза, называемого еще болезнью Шагаса (Т. cruzi), который распространяется кровососущими триатомовыми клопами.
Трипаносомы облигатно связаны с двумя хозяевами: у человека и млекопитающих они паразитируют в виде трипомастигот, а в организме беспозвоночных и культуре — в виде эпимастигот.
В крови человека nрипаносомы имеются в виде веретеновидного тела длиной 25-30 мкм, с одним жгутиком, закрепленным блефаробластом на заднем конце. Жгутик соединяется с пелликулой посредством ундулирующей мембраны, расположенной вдоль клетки, и заканчивается свободным концом на передней части паразита. Кроме центрально расположенного ядра, клетка содержит кинетопласт (хорошо окрашиваемое тельце с ДНК).
У эпимастигот отсутствует ундулирующая мембрана, жгутик короткий, кинетопласт расположен ближе к центру. Инцистирование для трипаносом нехарактерно.
Лейшманииделят на три патогенных для человека вида (L.tropica,L.donovani,L.braziliensis), которые являются возбудителями кожного («восточная язва», болезнь Боровского), висцерального и кожно-слизистого («кала-азар») лейшманиоза, передаваемого через укусы москитов родовPhlebotomus,Lutzomyia. Жизненный цикл лейшманий характеризуется облигатной сменой хозяев: позвоночного (лейшманиальная стадия, или амастиготы) и беспозвоночного (лептомонадная стадия, или промастиготы).
Амастиготы лейшманий (или тканевые формы, поскольку в этой форме лейшманий внутриклеточно паразитируют в организме человека и других млекопитающих) — безжгутиковые овальные клетки размером 2—5 мкм, имеющие ядро, кинетопласт, блефаропласт.
Промастиготы — веретеновидные клетки, с заостренным концом (длина 20—30 мкм, ширина 5—6 мкм), на котором блефаропластом закреплен один жгутик; в их протоплазме различают ядро и кинетопласт. При культивировании на искусственных средах лейшманий растут в промастиготной форме. Цист лейшманий не образуют.
Класс саркодовые— типичные амебовидные паразиты, передвигаются с помощью псевдоподий, размножаются бесполым путем, бинарным делением.
У человека паразитирует дизентерийная амеба(Entamoeba hystolytica), вызывающая амебную дизентерию, или амебиаз. Выделяют четыре морфологически и функционально различающиеся вегетативные формы: тканевую, большую вегетативную, просветную, предцистную.
Во внешней среде амебы находятся в виде четырехъядерных цист (9—12 мкм) с плотной двухконтурной оболочкой. При попа-
42
рыдании в тонкую кишку человека из цист образуется восемь одно-ядерных вегетативных форм, заселяющих толстую кишку.
Тканевая форма (20—25 мкм) имеет крупное ядро, обладает амебоидным движением, в цитоплазме различают два слоя — эндо- и эктоплазма. Инвазивные свойства амеб связаны с тканевыми формами.
Большая вегетативная форма является наиболее крупной, ее размеры достигают при вытянутых псевдоподиях 60—80 мкм. В эн-доплазме встречаются фагоцитированные эритроциты.
Просветная форма (15—20 мкм) обитает в просвете толстой кишки. Передвигается медленно, в цитоплазме выявляются фаго-цитированные бактерии.
Предцистная форма (12—20 мкм) отличается наиболее мед-ленными движениями, гомогенной цитоплазмой, отсутствием включений.
Класс споровики— паразиты со сложным жизненным циклом, включающим половое и бесполое размножение и смену хозяев, не имеют специальных органов движения, только некоторые формы на определенных стадиях развития образуют псевдоподии и жгутики. К этой группе относятся малярийные плазмодии и токсоплазмы.
Плазмодии малярии.Паразитами человека как промежуточного хозяина являются четыре вида плазмодиев малярии:Plasmodium vivax и Р. оvа1е — возбудители трехдневной малярии; Р.mаlariae — возбудитель четырехдневной малярии; Р.falciparum— возбудитель тропической малярии.
Все возбудители, независимо от вида, проходят в человеческом организме бесполую фазу развития (шизогония). Вторым, окончательным хозяином является самка малярийного комара рода Anopheles, в желудке которой происходит половая фаза развития |лазмодия (спорогония).
Бесполый цикл развития малярийного плазмодия соcтоит из двух последовательных процессов со сменой среды обитания паразита. При укусе зараженный комар со слюной вносит кровь человека малярийных паразитов в виде спорозоитов — одноядерных клеток серповидной формы, длиной 14 мкм.
Спорозоиты внедряются в клетки печени и проходят преэрит-юцитарный, или тканевой цикл развития, последовательно претерпевая стадии тканевого шизонта, в результате деления (меруляции) которого образуется множество тканевых мерозоитов. Последние проникают в эритроциты, и с этого момента начинается их развитие в крови — эритроцитарный цикл.
Мерозоиты в эритроцитах, питаясь гемоглобином, быстро растут, и через несколько часов превращаются в молодые шизонты, имеющие вид кольца («перстня с рубином»), поскольку при окраске по методу Романовского — Гимза центральную часть этой
43
округлой клетки занимает большая вакуоль, оттесняющая к периферии голубую цитоплазму и рубиново-красное ядро. Процесс роста плазмодия сопровождается увеличением его массы и уменьшением вакуоли; взрослый шизонт почти полностью заполняет эритроцит. Через 48—72 часа паразит делится путем множественного дробления, образуя мерозоиты.
Разрушая эритроцит, мерозоиты выходят в кровяное русло, часть их, не подвергшаяся фагоцитозу, внедряется в новые эритроциты, и эритроцитарный цикл шизогонии повторяется.
Процесс эритроцитарной шизогонии строго цикличен, его продолжительность у разных видов плазмодиев различна: Plasmodium vivax и Р. оvа1е, Р.falciparum— составляет 48 часов, для Р.mаlariae — 72 часа.
Приступ лихорадки при малярии совпадает с моментом разрушения эритроцитов и выхода мерозоитов, поскольку в кровь поступают токсические продукты жизнедеятельности паразита и распада эритроцитов.
Образование половых клеток (гаметогония). Во время эритроцитарных циклов некоторые мерозоиты проникают в эритроциты и дифференцируются на мужские и женские гаметоциты (предшественники половых клеток). Таким образом, половой цикл развития начинается в человеческом организме, но для его продолжения гаметоциты должны с кровью попасть в организм самки комара.
Половое размножение в организме комара (спорогония). При укусе вместе с кровью больного человека в желудок самки комара попадают половые (гаметоциты) и бесполые (мерозоиты) клетки малярийного плазмодия. Бесполые клетки погибают, а половые дозревают, превращаясь в микро- и макрогаметы, которые, сливаясь, образуют зиготу, прикрепляющуюся к стенке желудка комара. На одном из концов зигота заостряется и превращается в оокинету. Оокинеты благодаря значительной подвижности и заостренной форме проникают в толщу стенки желудка. Под наружным слоем желудка они превращаются в ооцис-ты. Внутри ооцисты происходит деление паразита на тысячи спорозоитов, которые после созревания с гемолимфой поступают в слюнные железы комара.
Морфологические особенности отдельных видов плазмодиев малярии. У Р. Vivax юный шизонт имеет форму правильного кольца, иногда в одном эритроците встречаются две-три особи; зрелые шизонты приобретают амебоидную форму; в стадии меруляции шизонт делится на 12—24 мерозоита.
Морфология Р. оvа1е сходна с предыдущим видом, но он образует 6—12 мерозоитов.
44
Зрелый шизонт Р. malariae имеет лентовидную форму; в процессе его меруляции образуется 6—12 мерозоитов, располагающихся в виде розетки. Р.falciparum дробится на 12—24 мерозоита, отличается морфологическими особенностями гематоцитов, имеющих полулунную форму.
Токсоплазмы —простейшие, инфицирующие различные виды
животных и птиц. У них наблюдается чередование полового и бесполого размножения со сменой хозяев. Окончательными хозяевами являются кошки и другие животные семейства кошачьих. Человек — один из промежуточных хозяев. У людей эти простейшие вызывают токсоплазмоз, который в основном протекает бессимптомно.
Морфологически токсоплазмы представляют собой серповидные клетки длиной 5—7 мкм и шириной 2—4 мкм, с центрально расположенным ядром. Благодаря своеобразной системе микротрубочек они совершают медленные вращательные и скользящие движения.
Токсоплазмы могут образовывать в мозге и некоторых других
тканях цисты (30—300 мкм), имеющие плотную оболочку и содержащие несколько тысяч паразитов.
Класс реснитчатые— простейшие, имеющие наиболее слож-ную внутреннюю структуру, в частности, несущие на поверхности клеток реснички, которые располагаются характерными участками, с двумя видами ядер — макро- и микронуклеусом.
Единственный представитель этой группы, паразитирующий в организме человека — кишечный балантидий (Ваlantidium coli), который имеет реснички и обитает в кишечнике человека и свиней. Его жизненный цикл состоит из двух фаз — бесполой (размножение поперечным делением) и половой (конъюгация). После конъюгации балантидий инцистируется. Цисты имеют овальную |форму (45—60 мкм) и покрыты плотной двухслойной оболочкой. Кишечный балантидий паразитирует в толстом кишечнике, вырывая балантидиаз, который является довольно редким заболеванием.
ЦАРСТВО ВИРУСЫ (У1КА)
Морфология и ультраструктура вирусов. Особенности классификации
Вирусы — это формы жизни, которые относят к отдельному царству Vira. Наука о вирусах — вирусология — одна из самых молодых медицинских наук.
В настоящее время вирусы определяют как самостоятельные
самопродуцирующиеся неклеточные структуры, способные фун-
45
кционировать в восприимчивых к ним клетках животных, растений, бактерий.
Целый ряд признаков отличает вирусы от прокариотов и эука-риотов:
отсутствие клеточной структуры — это неклеточные формы жизни;
наличие у вирусов только одной из двух нуклеиновых кислот, в то время как у всех остальных микроорганизмов имеются ДНК и РНК;
отсутствие собственных белоксинтезирующих систем. Синтез вирусных белков осуществляется белоксинтезирующим аппаратом клетки-хозяина, в которой вирус паразитирует;
уровень паразитизма у вирусов, в отличие от внутриклеточного паразитизма бактерий и простейших, определяется как генетический паразитизм;
вирусы не растут, и размножение у них происходит путем дисъюнктивной репродукции. В клетке отдельно синтезируются белки и нуклеиновые кислоты вирусов, а затем происходит их сборка в вирусные частицы.
В то же время вирусы, безусловно, обладают основными свойствами всех других форм жизни — способностью размножаться, наследственностью, изменчивостью, приспособляемостью к условиям внешней среды; они занимают определенную экологическую нишу, на них распространяются законы эволюции органического мира на земле.
Морфологию и ультраструктуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки у бактерий. Размеры вирусов определяют методом ультрафильтрации через фильтры с известным диаметром пор, а также методом ультрацентрифугирования. Одним из наиболее мелких вирусов является вирус полиомиелита, имеющий размеры 20—24 нм, а к наиболее крупным вирусам относятся вирусы оспы — около 350 нм.
Вне клетки-хозяина вирус существует в виде вириона. Форма вириона может быть различной — палочковидной, сферической, пулевидной (рис. 4). Центральную часть вириона занимает нуклеиновая кислота — ДНК или РНК, которая является хранителем наследственной информации, выполняя таким образом функции генома. У РНК-содержащих вирусов различают вирусы с «плюс»-нить РНК-геномом, поскольку РНК этих вирусов выполняет не только наследственную, но и плюс функцию информационной РНК. К «плюс»-нитевым, или вирусам с позитивным геномом относятся пикорнавирусы, тогавирусы, коронавирусы, ретрови-русы.
У РНК-содержащих вирусов с «минус»-нить геномом РНК выполняет только наследственную функцию. К вирусам с нега-
46
Iridovirus
Pox virus
![]()
Herpesvirus Adenovirus Papovaviridae Parvorirus ДНК-содержашие вирусы



Paramyxovirus Qrthomyxavirus Coronavius Arenavirus Leukovirus
![]()
Reovirus Picornaviridae Rhabdovirus Togaviridae Супергруппа
Буниамвера
Arboviruses
РНК-содержащие вирусы
Рис. 4. Форма и относительные размеры вирусов (по Ф. Феннеру и соавт., 1977)
тивным геномом относятся ортомиксовирусы, парамиксовирусы, буньявирусы, рабдовирусы. РНК этих вирусов не вызывает ин- фекционного процесса.
РНК-геном в основном является гаплоидным, но у ретрови- русов он диплоидный, так как состоит из двух однонитевых моле- кул РНК.
У ДНК-содержащих вирусов ДНК может быть представлена как однонитевыми, так и двунитевыми, линейными и кольцевы-
47
ми молекулами. В геномах, представленных двунитевыми ДНК, информация обычно закодирована на обеих нитях ДНК, что свидетельствует о максимальной экономии генетического материала у вирусов, как генетических паразитов.
Нуклеиновую кислоту вирусов окружает белковый капсид. Существуют два типа строения капсидов вирионов. В одном случае структурные единицы капсида — капсомеры ассоциируются с геномом и образуют спиралевидную винтообразную структуру. Такой тип укладки называется спиральным типом симметрии, а сама структура — нуклеокапсидом. В другом случае капсомеры образуют полое изометрическое тело, в центре которого находится геном. Такая укладка называется кубическим типом симметрии (см. рис. 4).
Сложно устроенные вирусы имеют липопротеидную оболочку — суперкапсид (см. вкл. III). Под оболочкой вириона подразумевается липогликопротеидный слой, который формируется в процессе почкования вируса на мембране клетки-хозяина.
Капсид и суперкапсид защищают вирионы от физических и химических воздействий и прежде всего от ферментов нуклеаз, а также обусловливают адсорбцию определенными клетками, определяют антигенные и иммуногенные свойства вирионов.
На основании типа и структуры нуклеиновой кислоты (РНК или ДНК), наличия или отсутствия липопротеидной оболочки вирусы делят на семейства. Все известные в настоящее время вирусы подразделяются на 19 семейств, из них 7 — ДНК- и 12 — РНК-содержащие вирусы. В основу деления вирусов на подсемейства положены следующие основные критерии: стратегия вирусного генома, размер, морфология вириона, тип симметрии, число капсомеров; круг восприимчивых хозяев; патологические изменения в клетках; географическое распространение, способ передачи и антигенные свойства. Вид вируса биноминального названия, как у бактерий, не получил. Однако на практике большинство вирусологов продолжает условно подразделять вирусы в соответствии с типом хозяина, поскольку вирусы поражают позвоночных и беспозвоночных животных, растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы участвуют также в процессах канцерогенеза, и могут приводить к развитию миокардитов, панкреатитов, иммунодефицитов и т. д.
Кроме обычных вирусов известны так называемые неканонические вирусы — прионы, которые представляют собой белковые инфекционные частицы, имеющие вид фибрилл размером 10—20х100—200 нм. Прионы, очевидно, являются одновременно индукторами и продуктами автономного гена человека или животных и в условиях медленной вирусной инфекции вызывают у них энцефалопатии.
48
Близкими к вирусам агентами являются вирусоподобные структуры — плазмиды и вироиды. Плазмиды, или эписомы, эпивирусы представляют собой две нити ДНК, образуемые клеткой, но не связанные с клеточной хромосомой. Плазмиды обеспечивают некоторые свойства бактерий, например, устойчивость к антибиотикам. Позднее плазмиды были обнаружены у эукариотов (грибов). Некоторые вирусы животных могут существовать в виде плазмид.
Вироиды представляют собой небольшие молекулы кольцевой суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Вирусы бактерий — бактериофаги
Вирусы, способные паразитировать в бактериальных клетках, репродуцироваться в них и вызывать их растворение (лизис), получили название бактериофаги, или просто фаги. В настоящее время эти вирусы выявлены не только у большинства болезнетворных бактерий, но и у некоторых грибов.
Большинство фагов под электронным микроскопом имеют форму головастика или сперматозоида (рис. 5), но могут быть нитевидной или кубической формы, размером от 20 до 800 нм. Наиболее хорошо изучены крупные бактериофаги, имеющие форму сперматозоида. Они состоят из икосаэдрической головки размером
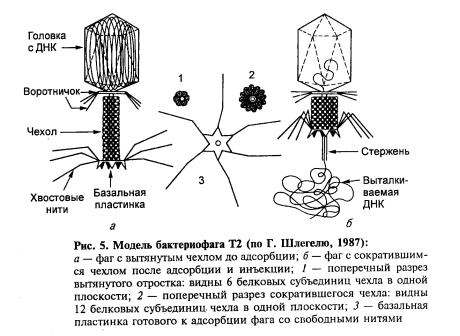
65—100 нм и хвостового отростка длиной 100 нм. В головке содержится нуклеиновая кислота — ДНК, реже РНК. Она окружена белковым капсидом.
Структурные белки фага различаются по составу полипептидов и представлены в виде множества идентичных субъединиц, уложенных по спиральному или кубическому типу симметрии. Кроме структурных белков, у некоторых фагов обнаружены внутренние (геномные) белки, связанные с нуклеиновой кислотой и белки-ферменты (лизоцим, АТФ-аза, участвующие во взаимодействии фага с клеткой). Внутри хвостового отростка имеется полый цилиндрический стержень, сообщающийся отверстием с головкой, снаружи — чехол, который способен сокращаться. Хвостовой отросток заканчивается шестиугольной базальной пластиной с короткими шипами, от которых отходят нитевидные структуры — фибриллы. У некоторых видов фагов чехол не может сокращаться. Фаги встречаются везде, где есть или были бактерии. Например, дизентерийные, брюшнотифозные фаги обнаруживаются в стоячей воде, почве, испражнениях человека.
Практическое применение фагов обусловлено их строгой специфичностью. Фаги используют для терапии и профилактики инфекционных заболеваний, а также при лабораторной диагностике для определения вида или штамма микроорганизмов. Препараты бактериофага выпускают в жидком виде и в виде таблеток.
ФИЗИОЛОГИЯ ПРОКАРИОТОВ
Предмет физиологии бактерий — исследование функций, то есть всех физических, химических и биологических процессов, происходящих в бактериальной клетке, а также физических, химических и биологических превращений, вызываемых микроорганизмом в окружающей среде.
ХИМИЧЕСКИЙ СОСТАВ БАКТЕРИАЛЬНОЙ КЛЕТКИ
Основой изучения физиологии бактерий является исследование химического состава этих микроорганизмов. Анатомия и химический состав бактерий являются рамками, а следовательно, и материалом, за счет которого развиваются жизненные процессы.
Химический состав прокариотов почти такой же, как и других живых организмов: состоит из двух компонентов — воды и сухого остатка, представленного смесью органических и минеральных соединений.
Основу микробной клетки составляет вода — 80—90 % общей массы. Вода находится в свободном и связанном состоянии.
50
Свободно содержащаяся в клетке вода необходима бактериям как растворитель органических и минеральных соединений; дисперсионная среда для коллоидов; источник водородных и гидроксильных ионов; фактор осмотического давления (тургор клетки).
С водой, как главным химическим компонентом структуры, связаны основные процессы жизнедеятельности бактериальной клетки — питание, дыхание, рост и размножение. Подчеркивая особую роль воды в определении химического состава и жизнедеятельности бактериальной клетки, необходимо обратить внимание на главную особенность — она должна быть биологически доступна для бактерий. Биологическая зона воды находится в температурном диапазоне от 2 °С (или ниже в растворах с высоким осмотическим давлением) до 100 °С.
Сухой остаток (10—20 % массы бактерий) представляет собой смесь органических и минеральных соединений, основу которых составляют четыре элемента (так называемые органогены) — азот, углерод, водород и кислород, присутствующие в различных сочетаниях в молекулах и свободном состоянии.
Минеральный состав бактерий характеризуют фосфор, сера, натрий, магний, калий, кальций и др.
Органические компоненты химического состава бактерий представляют белки, нуклеиновые кислоты, углеводы, липиды, витамины и др.
Более 50 % сухого остатка бактерий составляют белки, ответственные за жизнеопределяющие функции всех организмов, в том числе и микроорганизмов.
Различают простые и сложные белки бактерий.
Простые белки(протеины) при гидролизе распадаются до аминокислот, которые бактериальная клетка использует как источник углерода.
Сложные белкисостоят из протеина (простого белка) и нуклеиновой кислоты. Сложные белки наиболее важны для жизнедеятельности бактерий. Они выполняют пластическую и строительную функции; участвуют в процессе роста и размножения;
определяют видовые особенности бактерий; характеризуют антигенные и иммуногенные свойства; ответственны за наследственную передачу видовых признаков; обладают токсичностью и вирулентностью; в составе ферментов характеризуют биохимическую активность бактерий.
Нуклеиновые кислоты(10—30 % сухого остатка) представлены у бактерий двумя типами — ДНК и РНК. ДНК содержится в составе бактериальной хромосомы, РНК — в рибосомах, зернистых включениях. Биологическая роль молекулы ДНК связана с определением наследственных свойств бактерий. РНК (информационная, транспортная, рибосомальная) выполняет соответствую-
51
щие функции в информационной потребности клетки, в синтезе белков.
Углеводысоставляют 10—30 % сухого остатка, представлены в виде моносахаридов, дисахаридов и полисахаридов. Их функции:
пластическая;
энергетическая;
агрессивность, токсичность, аллергенность;
типовая специфичность;
питательная и запасная (гликоген, крахмал).
По содержанию азота бактериальные полисахариды подразделяются на азотсодержащие полисахариды, например, гексозамины (глюкоза + глюкозамин); безазотистые полисахариды (полимеры альдобионовой кислоты).
При полном гидролизе бактериальные полисахариды образуют глюкозу и глюкуроновую кислоту.
Липидыу большинства бактерий составляют 5—10 %, у дрож-жеподобных грибов и микобактерий достигают до 40 % сухого остатка.
В составе микроорганизмов липиды встречаются в виде простых жиров (глицерин и высшие кислоты) и сложных липидов (фосфолипиды).
Значительная часть липидов находится в комплексной связи с белками и углеводами. Они являются необходимыми компонентами цитоплазматической мембраны и клеточной стенки и выполняют роль запасных питательных веществ; энергетического материала; фактора устойчивости микроорганизмов к действию внешней среды (спора, клеточная стенка микобактерий).
Минеральный составмикроорганизмов представлен большей частью элементов таблицы Менделеева. Минеральные вещества входят в состав различных клеточных структур бактерий. Общее их содержание в расчете на зольный остаток после сжигания бактерий составляет 2—30 % и зависит от вида и той питательной среды, в которой выращивались бактерии.
В составе золы бактерий преимущественно определяются окиси металлов - Р205, (9-50 %), Nа20 (11-33 %), К2О (7-25 %), Мg0 (0,1-9 %), СаО (7-12 %), в виде неорганических примесей — также Si, Сl, Аl, Сu, Мnи др.
Основное назначение минералов
1. Регуляторы осмотического давления, рН, окислительно-восстановительного потенциала.
2. Катализаторы активности бактериальных ферментов.
3. Обязательная составная часть главных органоидов бактериальной клетки, ответственных за ее жизнедеятельность и жизнеспособность, так:
P - составная часть нуклеиновых кислот;
Fе — компонент цитохромоксидазы, каталазы, пероксидазы;
52
Сu— составляющая дыхательных ферментов;
S— неорганический компонент белков и аминокислот.
Ростовые вещества — факторы роста, биоактиваторы — важнейшие регуляторы обменных процессов, роста и размножения микроорганизмов. В зависимости от фактора роста микроорганизмы подразделяются на:
прототрофы— способные синтезировать все необходимые для Сроста и размножения органические соединения;
ауксотрофы— их рост и размножение невозможны при от-cутствии вещества, которое бактериальная клетка не способна или утратила способность синтезировать.
Непременным условием роста и размножения ауксотрофов является обязательное присутствие в среде обитания необходимого вещества. К факторам роста относятся, прежде всего, витамины (В1— тиамин, В2— рибофлавин, В6— пиридоксин, Н — биотин, В12— цианкобаламин, РР — никотиновая кислота, витамин К1— филлохинон). Ауксотрофы нуждаются также в поступлении извне определенных аминокислот. Для каждого представителя ауксо-трофов такая аминокислота строго определенная и незаменимая. Потребность микроорганизмов в факторах роста не является постоянной и может изменяться в зависимости от:
условий культивирования.Так, плесневый гриб Мuсоr rouuuxii нуждается в витаминах В1, В6лишь при росте в анаэробных усло-виях, а в аэробных условиях эти витамины он синтезирует самостоятельно;
химического состава окружающей среды.Большинство вита-минов входит в состав коферментов, поэтому микроорганизмы могут обойтись без необходимых витаминов, если продукты соот-ветствующей ферментативной реакции содержатся в самой среде.
Таким образом, потребность в витаминах выражает иногда потребность в недостающем ферменте или продуктах его актив-кности.
Пример. Возбудитель дифтерии нуждается в пантотеновой кислоте. Одновременно известно, что пантотеновая кислота синтезируется из пантоевой кислоты и β-аланина. Поэтому потребность дифтерийной палочки в пантотеновой кислоте можно удов-летворить, внеся в питательную среду β-аланин.
ПИТАНИЕ БАКТЕРИЙ
Для роста и размножения микроорганизмов необходимы источники питания. Питание обеспечивает бактериальную клет|ку пластическим материалом для самовоспроизведения и энергией. Однако при наличии необходимых источников энергии и питания скорость размножения микроорганизмов в значительной мере
53
зависит от условий среды. Поэтому механизмы клеточной саморегуляции можно условно разделить на две основные группы: неспецифические механизмы регуляции роста и размножения; специфические механизмы саморегуляции.
К группе неспецифическихмеханизмов относится совокупность действия различных физико-химических факторов, регулирующих общую скорость всех основных процессов жизнедеятельности:
температура, рН, концентрация ионов, степень обеспечения среды кислородом и т. д.
Таким образом, эту форму регуляции можно определить еще как неконтролируемую со стороны клетки,то есть характеризующую действие факторов внешней среды.
Специфическиемеханизмы обменных процессов бактериальной клетки возникли из неспецифических и обязательно включают их в себя, однако, представляют более сложную форму взаимосвязи.
Основу специфической саморегуляции живых систем составляет ферментсубстратная реакция.
Бактериальная клетка имеет обширный спектр ферментов. Например, в геноме Е.соli штамм К-128 содержится информация, достаточная для кодирования около 4000 индивидуальных белков.
Способы питания микроорганизмов определяются в зависимости от источника энергии; донора водорода (электронов); источника углерода.
Источник энергии.По этому показателю отмечают два главных типа метаболизма:
1. Фототрофные бактерии, использующие в качестве источника энергии для роста электромагнитное излучение (свет). Среди них различают:
анаэробные фототрофные бактерии, не выделяющие молекулярный кислород;
аэробные фототрофные бактерии (водоросли), выделяющие на свету кислород.
2. Хемотрофные (хемосинтезирующие) бактерии, получающие энергию в результате окислительно-восстановительных реакций с участием субстратов, которые служат для них источником питания.
Доноры водорода.По этому критерию бактерии подразделяются на:
метатрофные, использующие в качестве доноров электронов неорганические элементы (H2, NH3, НS, S, СО,Fe2+ и др.);
органотрофные, для которых донорами водорода (электронов) служат органические соединения.
Источники углерода.По этому показателю бактерии подразделяются на:
54
аутотрофные, получающие углерод путем фиксации СО2из воздуха;
гетеротрофные, которые усваивают углерод из органических соединений.
Гетеротрофы делятся на сапрофитов и паразитов.
Сапрофиты (от лат. saprophyticus) — произрастающие за счет мертвых субстратов.
Паразиты — живущие за счет органических соединений человека и животного. Именно к ним относятся возбудители инфекционных заболеваний человека и животных. Признак паразитизма может быть абсолютным (облигатным) и относительным (условная патогенность).
Механизмы питания.Первооснову механизма питания бактерий составляет разница осмотического давления, основанная на различии концентраций между питательной средой и цитоплазматическим составом бактериальной клетки. Принципиальное значение для понимания механизмов питания имеет то, что их внутриклеточное осмотическое давление, как минимум, в два раза ниже, чем у клетки животного происхождения.
Согласно законам физической химии, при разнице осмотического давления жидкость движется в сторону более высокой концентрации, а растворенное в ней вещество — в сторону меньшей концентрации.
Молекула растворенного вещества может пройти через ци-топлазматическую мембрану, если на нее действует какая-нибудь сила или существуют механизмы, которые обеспечивают перенос этой молекулы через мембрану. Поэтому различают следующие механизмы питания.
1. Пассивная диффузия —перенос питательных веществ в клетку за счет разности концентраций по обе стороны цитоплазматической мембраны. Наблюдается в основном при поступлении в бактериальную клетку воды. Другие молекулы за счет пассивной диффузии практически не попадают.
2. Облегченная диффузия(перенос «по течению») — прохождение большинства растворенных веществ через цитоплазматическую мембрану осуществляется за счет их захвата на внешней и переноса к внутренней поверхности мембраны молекулами-переносчиками, которые называются пермеазами. Они характеризуются разной степенью специфичности к веществам.
3. Активный перенос —против градиента концентрации, то есть «против течения», осуществляется со значительным расходованием энергии АТФ, выделяемой в процессе метаболизма.
Пример. Е.соli активно поглощает О2. Прохождение одной молекулы кислорода через цитоплазматическую мембрану связано с расходом одной молекулы АТФ.
55
4. Транслокация радикалов —перенос химически измененных молекул, которые в целом виде не способны пройти через ци-топлазматическую мембрану. Экзоферменты фрагментируют их до единичных радикалов, каждый из которых по отдельности пермеазы переносят через цитоплазматическую мембрану внутрь клетки, где происходит ресинтез этого соединения.
ФЕРМЕНТЫ БАКТЕРИЙ
Ферментные системы бактериальной клетки условно классифицируются на следующие основные группы:
системы, обеспечивающие активный транспорт питательных веществ в клетку из окружающей среды,— пермеазы;
системы, регулирующие превращение энергии в клетке;
системы, осуществляющие биосинтез аминокислот и нуклеотидов, то есть предшественников белков и нуклеиновых кислот;
системы, обеспечивающие собственно синтез белков и нуклеиновых кислот;
системы, регулирующие репликацию и сегрегацию хромосом, то есть деление ядерного аппарата;
системы, регулирующие собственно клеточное деление;
системы биосинтеза липидов, полисахаридов и клеточных мембран.
Согласованное взаимодействие этих систем и составляет сущность процессов клеточной саморегуляции. Решающее значение при этом имеют:
согласованность скоростей реакций;
строгое соблюдение последовательности их включения;
регулирование количественного и качественного состава ферментов.
Конечным результатом регуляции биосинтетических процессов в клетке является воспроизведение потомства, а показателем сбалансированности — скорость роста бактерий.
По характеру контролируемых реакций бактериальные ферменты подразделяются на такие основные группы:
гидролазы,осуществляющие гидролиз (эстеразы, протеазы, нуклеазы);
трансферазы,обеспечивающие катализ путем переноса определенных радикалов от одной молекулы к другой;
окислительные ферменты(оксидоредуктазы), катализирующие процессы окисления — восстановления (оксидазы, пероксидазы, каталазы);
изомеразыирецемазы,играющие роль в углеводном обмене;
лиазы,обеспечивающие ферментацию негидролитическим путем определенных химических групп;
лигазы,регулирующие биосинтетические реакции клетки.
56
Наряду с ферментами обмена патогенные бактерии имеют ферменты агрессии, определяющие факторы их вирулентности: гиа-луронидазу, коагулазу, коллагеназу, нейраминидазу, дезоксири-бонуклеазу.
В зависимости от постоянства присутствия в бактериальной клетке и обязательности участия в ее клеточном обмене ферменты подразделяются на:
конститутивные,которые постоянно находятся в клетке независимо от условий ее существования,— это основные ферменты клеточного обмена (липазы, протеиназы и др.);
адаптивные(индуцибельные) — синтезируются только при наличии в среде соответствующего субстрата.
В зависимости от точки приложения действия бактериальные ферменты подразделяются на экзоферменты и эндоферменты. Экзоферментывыделяются клеткой во внешнюю среду, где производят расщепление сложных соединений (белков, жиров, углеводов) до более простых, доступных усвоению клеткой.
Эндоферментысодержатся внутри клетки, осуществляют пластическую функцию и синтез сложных соединений в клетке.
Поскольку ферменты бактерий обладают высокой специфичностью, именно это свойство широко используется для идентификации микроорганизмов.
ДЫХАНИЕ БАКТЕРИЙ
Необходимую для жизнедеятельности энергию бактериальная клетка получает за счет экзотермических химических реакций окисления различных химических соединений, обладающих запасами потенциальной энергии.
Сущность процесса дыхания бактерий заключается в протекании биохимических реакций, в результате которых образуется АТФ,являющаяся универсальным переносчиком химической энергии между взаимопротивоположными процессами выделения и потребления энергии.
Таким образом, под термином «дыхание» подразумевается окисление органических веществ клетки кислородом, в результате чего образуется конечный продукт — углекислый газ.
Одни бактерии для биохимических реакций окисления используют свободный кислород. Такой тип дыхания называется аэробным.Микроорганизмы, осуществляющие аэробный тип дыхания при наличии в атмосфере не менее 20 % кислорода, называются облигатными аэробами. 1
Другие бактерии получают энергию в результате окисленця, при котором в качестве акцепторов Н2-электронов выступают неорганические соединения. Такой процесс называется анаэробным
57
дыханием. Различают два основных типа анаэробного дыхания:
нитратный и сульфатный.
При нитратном типе источником энергии является восстановление нитратов до азота и аммиака.
Бактерии, использующие нитратный тип анаэробного дыхания, относятся к факультативным анаэробам.Среди них: кишечная, дизентерийная, брюшнотифозная палочки, стафилококки, стрептококки и др.
При сульфатном типе в результате окисления соединений образуется сульфат, который восстанавливается до сероводорода. Бактерии, использующие сульфатный тип дыхания, относятся к облигатным анаэробам,не способным расти в присутствии кислорода, поскольку при аэробном дыхании водородные атомы закономерно связываются с кислородом, в результате чего образуется чрезвычайно токсичная для бактериальной клетки перекись водорода (Н^О,), которая инактивируется двумя основными ферментами — каталазой и пероксидазой.
Облигатные анаэробы не содержат каталазы. Поэтому при наличии кислорода образующаяся перекись водорода оказывает губительное действие на облигатный анаэроб. Среди облигат-ных анаэробов — возбудители столбняка, газовой гангрены, ботулизма.
Особую группу по типу дыхания составляют микроаэрофилы. При необходимости использования кислорода микроаэрофилы отличаются от облигатных аэробов тем, что кислород им необходим в значительно меньшем количестве. К микроаэрофилам относятся актиномицеты, бруцеллы, лептоспиры.
РОСТ И РАЗМНОЖЕНИЕ БАКТЕРИЙ
По определению Э. Роуза, «рост — это координированная репликация (воспроизведение) всех структур, органелл и компонентов клетки микроорганизма».
Иными словами, под ростом бактериальной клетки следует понимать увеличение массы ее цитоплазмы, которое происходит в результате синтеза клеточного материала в процессе питания. На рис. 6 представлена типичная кривая роста популяции бактерий, в которой различают четыре стадии: лаг-фаза; экспоненциальная, или логарифмическая фаза; стационарная фаза; фаза отмирания.
Лаг-фаза(4—5 часов) наступает после того, как в среду внесен посевной материал. Это период адаптации бактерий к питательной среде, когда происходит дифференциальная активация экзо-и эндоферментов для последующего осуществления ферментсуб-стратной реакции. При стабильном содержании ДНК отмечается резкое повышение бактериального белка и РНК.
58
Рис. 6. Фазы роста популяции бактерий (по А. А. Воробьеву и соавт., 1994)
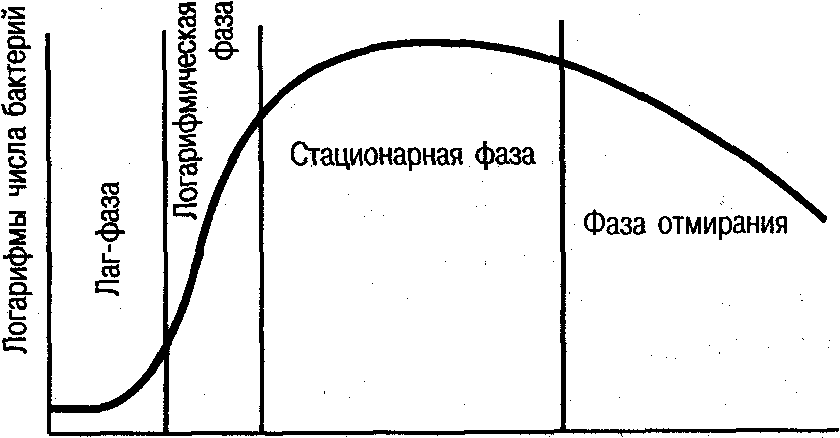
время
Длительность лаг-фазы, как правило, непродолжительна, измеряется часами и зависит от вида бактерий; кратности посева на данную среду; состояния культуры; температуры, используемой для выращивания; состава питательной среды.
При отсутствии видимых проявлений роста в лаг-фазе происходит увеличение биомассы, в результате чего размер бактериальной клетки возрастает в несколько раз.
Достигнув определенного размера, «накопив» нужное количество белка, РНК и ДНК, активировав экзо- и эндоферменты, бактериальная клетка начинает активно делиться. Размножение бактерий происходит путем поперечного деления клетки (см. вкл. III).
С началом деления наступает следующая стадия — фаза логарифмического роста(5—6 часов) — это фаза размножения, осу-ществляемая посредством бинарного деления материнской клетки на две дочерние. «Цепная» реакция прогрессивно ускоряющегося бинарного деления бактериальных клеток приводит к быстрому нарастанию бактериальной массы в питательной среде, интенсивному расходованию ее энергетического субстрата и накоплению продуктов бактериального метаболизма. В результате среда становится все более неблагоприятной для дальнейшего роста и размножения бактерий.
В этот период наступает третья, стационарная, фаза роста,во время которой скорость размножения остается постоянной. В зависимости от вида культивируемых бактерий может длиться долго, после чего наступает четвертая стадия — фаза отмирания,характеризующаяся прогрессивным отмиранием бактериальных клеток по логарифмическому типу. Продолжительность этой фазы — от 48 часов до нескольких недель.
Характер роста бактерий на жидких питательных средах различен:
диффузное помутнение питательной среды;
образование пленки или осадка («придонный рост»);
рост в виде «комочка ваты».
59
Характер роста на жидкой питательной среде используется для дифференциации бактерий.
Для культивирования бактерий в лабораторных условиях применяют искусственные питательные среды различного состава.
Обычные, или простые питательные среды (мясо-пептонный агар, мясо-пептонный бульон) используются для культивирования многих видов бактерий и приготовления сложных питательных сред.
К сложным относятся элективные и дифференциально-диагностические питательные среды. Элективные среды обеспечивают развитие определенного вида микроорганизма, при этом сопутствующая микрофлора не вырастает или растет очень медленно. Дифференциально-диагностические среды используются для изучения биохимических свойств микроорганизмов и дают возможность их дифференцировки по ферментативной активности.
По консистенции питательные среды могут быть жидкие;
полужидкие (0,3-0,7 % агара); плотные (1,5-2 % агара).