

Рестрикционный анализ молекул днк
Огромные возможности рестриктаз можно проиллюстрировать на следующем примере. Рассмотрим двухцепочечную репликативную форму бактериофага фХ174. Он содержит два ковалентно связанных комплементарных кольца, из 5386 нуклеотидов каждое. Чистый препарат ДНК состоит из гомогенных молекул фага. Теперь представим себе, что этот препарат подвергается действию эндонуклеазы, однократно разрезающей двойную спираль кольца без какой-либо специфичности в отношении точки разреза. В результате мы получим препарат ДНК, содержащий линейные молекулы 5386 различных типов, т. е. препарат, совершенно бесполезный с точки зрения анализа нуклеотидных последовательностей. Напротив, если при этом используется рестриктазаPst I, разрезающая палиндромную последовательность GTGCAG (выписана последовательность нуклеотидов лишь в одной цепи двойной спирали), то получается гомогенный препарат линейных молекул ДНК длиной до 5386 нуклеотидов каждая, упорядоченных в одной и той же последовательности. ГеномфХ174 содержит лишь один сайт, узнаваемый рестриктазой Pst I. В геноме фХ174 есть сайты, узнаваемые многими другими ферментами, перечисленными в табл. 9.1. Количество и локализация сайтов для каждой рестриктазы строго определены. Таким образом, воздействие каким-то ферментом приводит к образованию уже известного количества фрагментов ДНК фиксированного размера. Размер каждого типа фрагментов можно узнать с помощью электрофореза в геле: мелкие фрагменты перемещаются в геле быстрее крупных. Так как фрагменты каждого типа характеризуются одинаковым размером и одинаковой последовательностью нуклеотидов, то нуклеотидную последовательность в каждом из них можно определять отдельно, на выделенном посредством электрофореза в геле препарате. Полную последовательность нуклеотидов в геноме можно затем «собрать» из последовательностей отдельных фрагментов, если знать последовательность самих фрагментов в геноме.
9. Методы работы с ДНК271
|
Рис. 9.6. Электрофоретический анализ рестрикционных фрагментов ДНК. ДНК фага λ инкубировали с различными указанными на рисунке рестриктазами время, достаточное для того, чтобы во всех чувствительных сайтах произошло расщепление нуклеотидной последовательности. Образовавшуюся смесь фрагментов ДНК подвергали электрофорезу в агарозном геле. Полосы идентифицировали в ультрафиолетовом свете после окрашивания геля бромистым этидием. Стартовые точки обозначены жирными стрелками. Интактная ДНК фага λ представляет собой линейную молекулу длиной около 48 500 н. п. При действии рестриктазы BglII возникают фрагменты длиной 22800, 13600, 9800, 2300 н. п. (они отмечены тонкими стрелками) и 460 н. п. (неразличим). |
|

Геном фага фХ174 принадлежит к числу самых мелких и наиболее хорошо изученных (см. гл. 7), и именно для этого генома в 1978 году была полностью определена последовательность нуклеотидов с помощью описанного выше подхода (см. гл. 12).
Фаг λ также представляет собой хороший пример того, как можно использовать фрагменты, образующиеся при рестрикции (рестрикты) для описания структуры генома вируса. На рис. 9.6 можно видеть число и размеры фрагментов ДНК, образующихся при действии нескольких различных рестриктаз на геном этого фага. Последовательность фрагментов, образующихся при действии определенной рестриктазы, можно определить с помощью сочетания нескольких методов, цель которых состоит в построении карты сайтов рестрикции генома фага λ. На рис. 9.7 схематически изображена карта сайтов Eco RI и Hin dIII на фоне генетической и физической карт генома фага λ.
Последовательность образующихся при рестрикции фрагментов в интактной молекуле ДНК можно установить несколькими способами. Прежде всего можно использовать неполное расщепление ДНК эндонуклеазами и последующее разделение фрагментов электрофорезом. Затем
272Организация и передача генетического материала
|
Рис. 9.7. А. Физическая карта ДНК фага λ, на которой указаны сайты рестрикции для Eco Kl и Hin dIII. Б. Размеры фрагментов, образующихся при действии (1) Hin dIII; (2) Eco RI и (3) Hin dIII и Eco RI. [Murray К., Murray Ν. (1975). J. Mol. Biol., 98, 551.] |
|
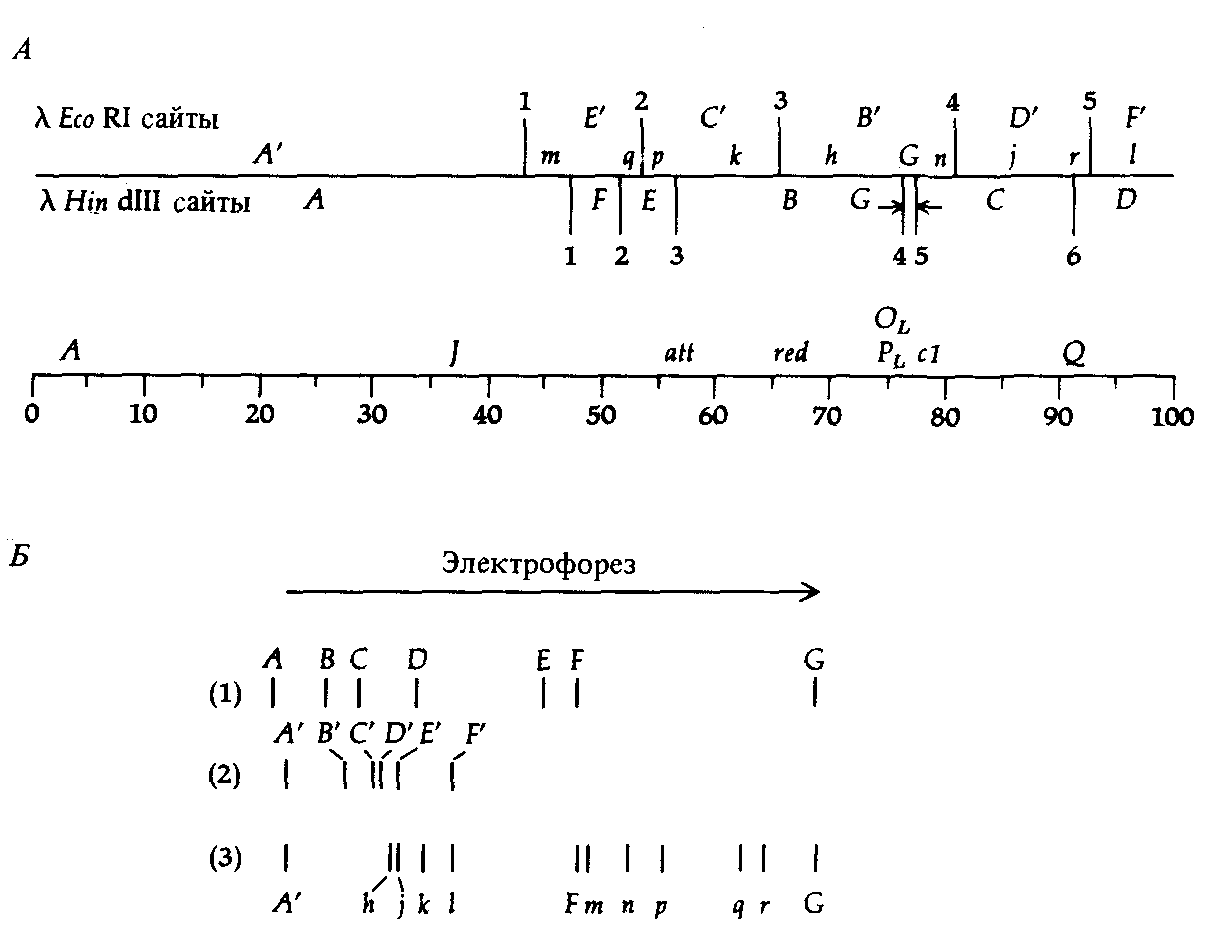
выделенные из электрофоретического геля крупные фрагменты снова подвергают действию того же фермента и посредством электрофореза устанавливают идентичность образующихся мелких фрагментов. С другой стороны, фрагменты, возникшие в результате полного расщепления под действием одного фермента, можно извлечь из геля, обработать второй рестриктазой и определить затем с помощью электрофореза размеры образовавшихся мелких фрагментов. Так, например, из рис. 9.7 видно, что выделение фрагмента А под действием рестриктазы Hin dIII и последующая обработка Eco RI приводят к образованию фрагментов А' и т, возникающих и при одновременном воздействии обоими ферментами.
Сопоставление карты рестрикции с генетической картой можно осуществить, действуя рестриктазами на ДНК, выделенную из различных мутантов фага λ с известными делециями и перестройками в геноме. Сравнивая фрагменты ДНК фагов дикого типа и мутантных, можно определять участки генетической карты фага λ, в которой локализованы соответствующие фрагменты.
Построение рестрикционной карты генома дает возможность разработать стратегию определения последовательности нуклеотидов в генах, представляющих особый интерес. В результате действия нескольких различных ферментов образуются сравнительно мелкие перекрывающиеся фрагменты, содержащие не более нескольких сотен нуклеотидов. Эти фрагменты могут быть выделены в чистом виде, и в них может быть установлена последовательность нуклеотидов. Затем, зная взаимно перекрывающиеся участки последовательностей, можно восстановить последовательность нуклеотидов в крупных фрагментах и в геноме в целом.
9. Методы работы с ДНК 273