

Сцепленное с полом наследование у человека и других видов
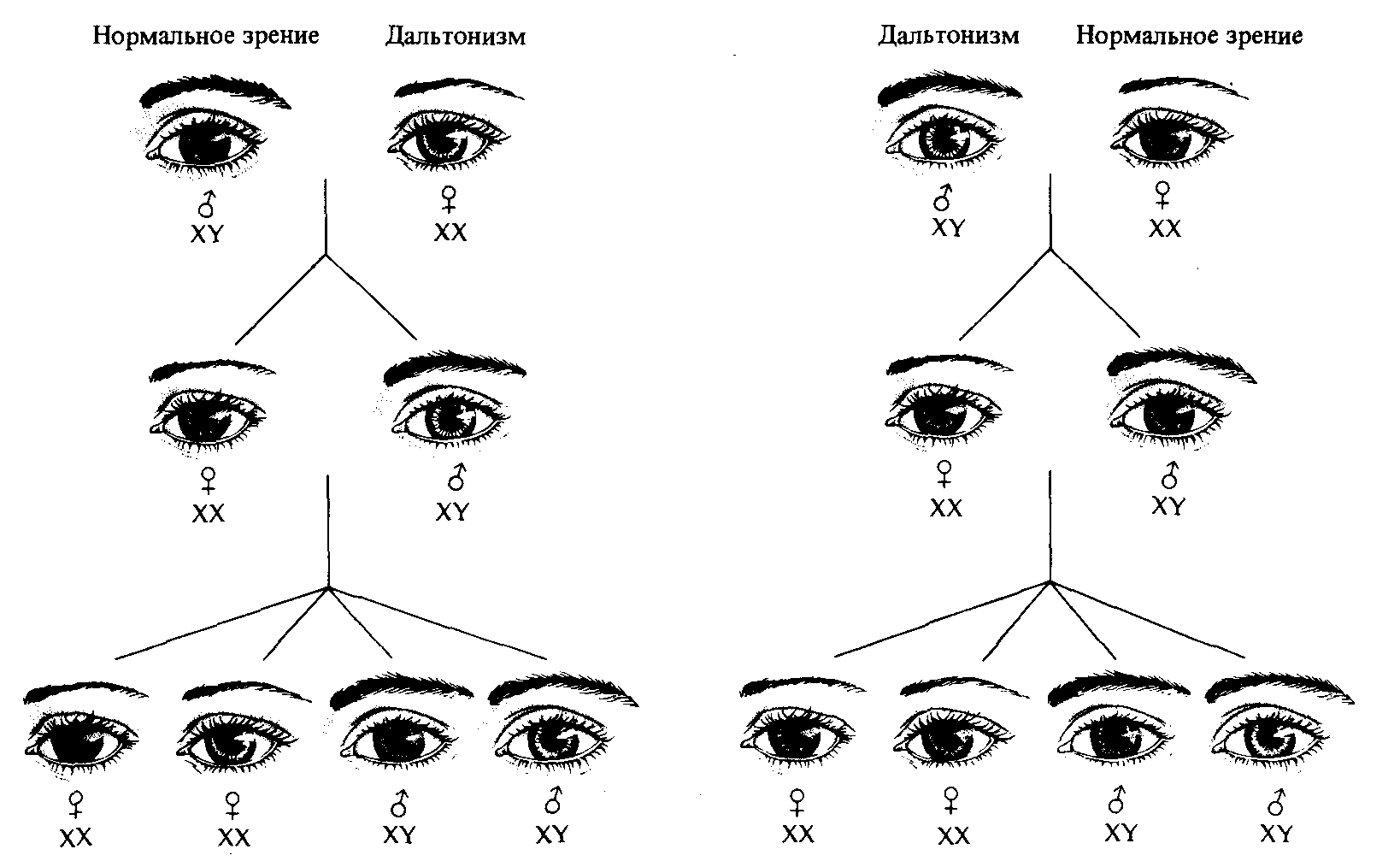
Механизм сцепленного с полом наследования, описанный выше для дрозофилы, характерен также для всех животных и растений, у которых мужской пол является гетерогаметным. Самцы называются гемизиготными по генам, расположенным в Х-хромосоме, поскольку в отношении этих генов они не являются ни гомо-, ни гетерозиготными. У человека известно около 150 признаков, сцепленных с полом. Характер наследования одной из форм дальтонизма -от матери к ее сыновьям - известен уже сотни лет. В 1911 году Эдмунд Вильсон указал на то, что все известные факты наследования дальтонизма можно объяснить, предположив, что дальтонизм определяется рецессивным сцепленным с полом аллелем, а мужской пол у людей гетерогаметен (рис. 3.10).
Другим примером сцепленного с полом признака у человека может служить гемофилия - тяжелое заболевание, для которого характерна неспособность крови свертываться. У нормальных людей при небольшом
|
|
|
Рис. 3.10. Наследование одной из форм дальтонизма у людей. Все сыновья женщины-дальтоника -дальтоники, все ее дочери-носители (гетерозиготы) гена дальтонизма. Мужчина-дальтоник передает ген дальтонизма всем своим дочерям, и через них он наследуется половиной сыновей. (Цвет глаз при этом, конечно, не играет роли; на рисунке неокрашенность радужной оболочки означает дальтонизм.) |
76 Организация и передача генетического материала
|
|
|
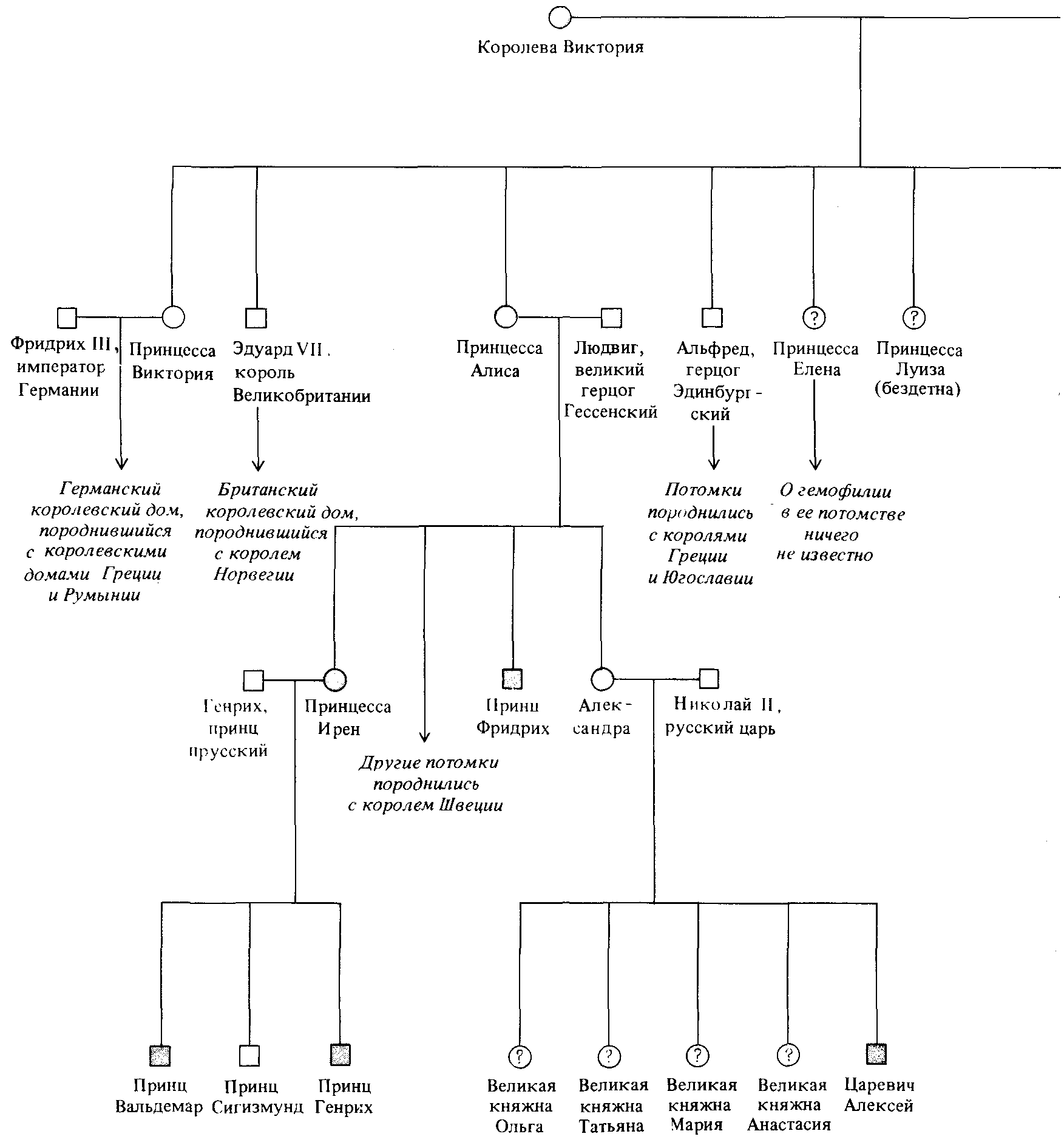
Рис. 3.11. Наследование гемофилии в потомстве королевы Виктории. Как это принято при построении родословных у человека, женщины изображены кружками, мужчины - квадратиками. Горизонтальные линии между квадратом и кружком означают супружество, вертикальные линии проведены к потомству. |
3. Хромосомные основы наследственности 77
|
|
78 Организация и передача генетического материала
|
|
|
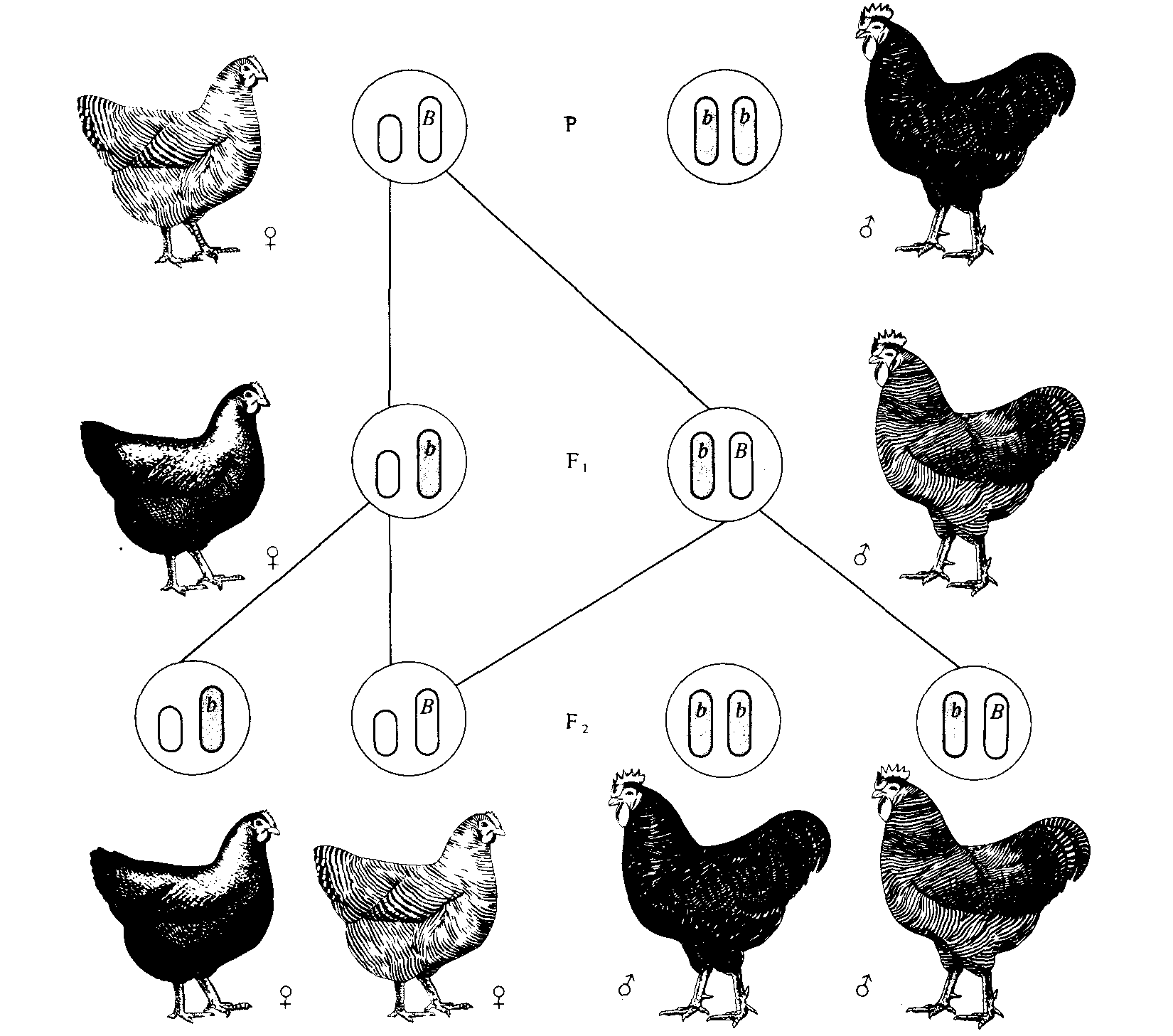
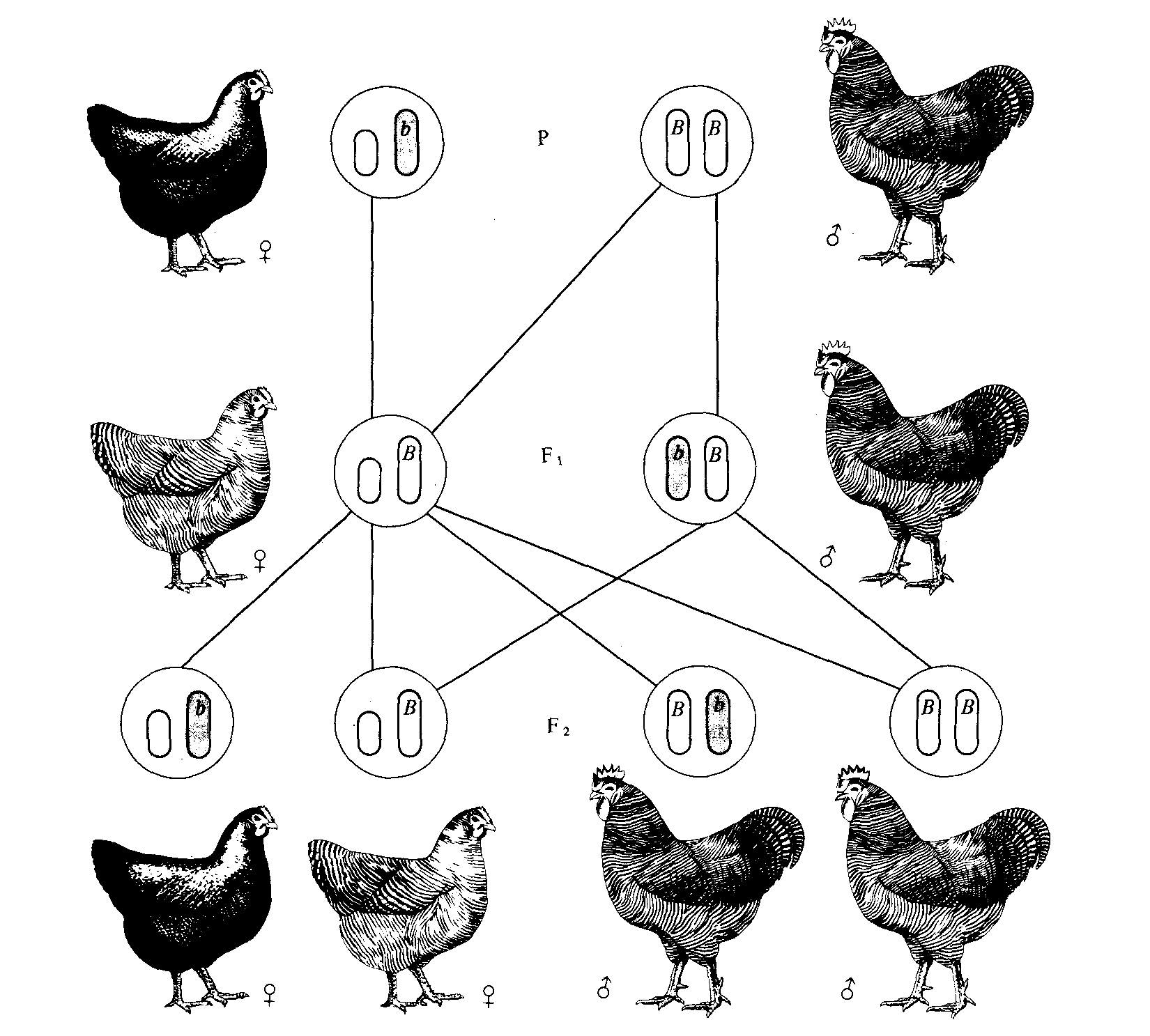
Рис. 3.12. Сцепленное с полом наследование у кур; скрещивание рябой курицы с гладким петухом. Ген рябой окраски (В) доминантен по отношению к гладкой окраске (b). У кур самки гетерогаметны, следовательно, куры передают Х-хромосому только сыновьям, а дочери получают Х-хромосому от отца. В результате дочери имеют рецессивный фенотип, унаследованный от отца, а сыновья — доминантный фенотип матери. Этот тип наследования отличается от изображенного на рис. 3.5, при котором доминантный фенотип матери наследуется всеми сыновьями и дочерьми. |
повреждении тканей кровотечение довольно быстро останавливается вследствие образования сгустков крови. Существуют по меньшей мере три типа гемофилии, два из которых обусловлены рецессивными сцепленными с полом генами, а один -очень редкий - определяется рецессивным аутосомным геном. Каждый из этих генов имеет отношение к одному из факторов, необходимых для нормального свертывания крови. Знаменитый пример сцепленного с полом наследования гемофилии представляет собой родословное древо некоторых правящих династий Европы, восходящее к королеве Виктории (рис. 3.11). Поскольку среди ее предков случаи гемофилии неизвестны, то естественно предположить,
3. Хромосомные основы наследственности79
|
|
|
Рис. 3.13. Сцепленное с полом наследование у кур: скрещивание гладкой курицы с рябым петухом. Все сыновья и дочери наследуют доминантный фенотип отца. Характер наследования отличается от изображенного на рис. 3.6, при котором доминантный фенотип отца проявлялся только у дочерей. |
что аллель, ответственный за гемофилию, возник в результате мутации в гамете одного из ее родителей.
Характер наследования сцепленных с полом признаков, описанный для дрозофилы и человека, меняется на противоположный, когда гетерогаметным полом являются самки. Например, у птиц, бабочек и некоторых рыб именно самки гемизиготны по сцепленным с полом признакам, и они передают гены, определяющие такие признаки, только своим сыновьям, тогда как самцы передают соответствующие гены как сыновьям, так и дочерям (рис. 3.12 и 3.13).
80 Организация и передача генетического материала
Y-хромосома
Обсуждавшиеся до сих пор гены, сцепленные с полом, расположены в Х-хромосоме и не имеют гомологичных аллелей в Y-хромосоме. Поэтому самцы (точнее, гетерогаметный пол) гемизиготны по сцепленным с полом генам, и эти гены всегда проявляются в их фенотипе, даже если они рецессивны.
Содержатся ли вообще какие-то гены в Y-хромосоме? Большинство генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако определенную генетическую информацию она все-таки несет. Различают два типа такой информации: во-первых, содержащуюся в генах, присутствующих только вY-хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так и в Х-хромосоме.
У видов, самцы которых гетерогаметны, Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно, для генов, содержащихся только в Y-хромосоме, характерно голандрическое наследование, т.е. они передаются только от отца к сыну и проявляются у самцов.
У человека наличие Y-хромосомы-необходимое условие принадлежности к мужскому полу: организмы, обладающие одной Х-хромосомой в отсутствие Y-хромосомы, характеризуются женским фенотипом, хотя большинство из них стерильны (этот генотип определяет так называемый синдром Тернера; см. гл. 21). Вообще, по-видимому, присутствие Y-хромосомы необходимо у животных для возникновения мужских половых признаков (или женских, если гетерогаметным поломявляются самки) или, во всяком случае, для плодовитости. У D. melanogaster особи с одной Х-хромосомой в отсутствиеY-хромосомыфенотипически выглядят как самцы, однако они стерильны: содержащиеся в Y-хромосоме гены необходимы для сперматогенеза.
У D. melanogaster известны гены, представленные как в Y-, так и в Х-хромосоме. Носители рецессивной мутации bobbed(bb) в гомозиготном состоянии характеризуются более короткими и тонкими щетинками, чем мухи дикого типа. Ген расположен в ядрышковом организаторе, т.е. в участке хромосом, ответственном за формирование ядрышка в интерфазе клеточного деления. При скрещивании самок, гомозиготных по рецессивному аллелю, с гетерозиготными самцами наблюдается необычное расщепление. Если носителем доминантного аллеля служит Х-хромосома самца, то все самки в F1 нормальны, а самцы обладают мутантным фенотипом (ХbbYbb). Если же носителем доминантного аллеля является Y-хромосома гетерозиготного отца (XbbYbb + ) то в потомстве F1, напротив, все самки имеют мутантный фенотип (ХbbХbb), а самцы-нормальный (XbbYbb +).
У человека в Y-хромосоме содержатся по крайней мере три гена, один из которых необходим для дифференциации семенников, второй требуется для проявления антигена гистосовместимости, а третий оказывает влияние на размер зубов.