

Нерасхождение х-хромосом
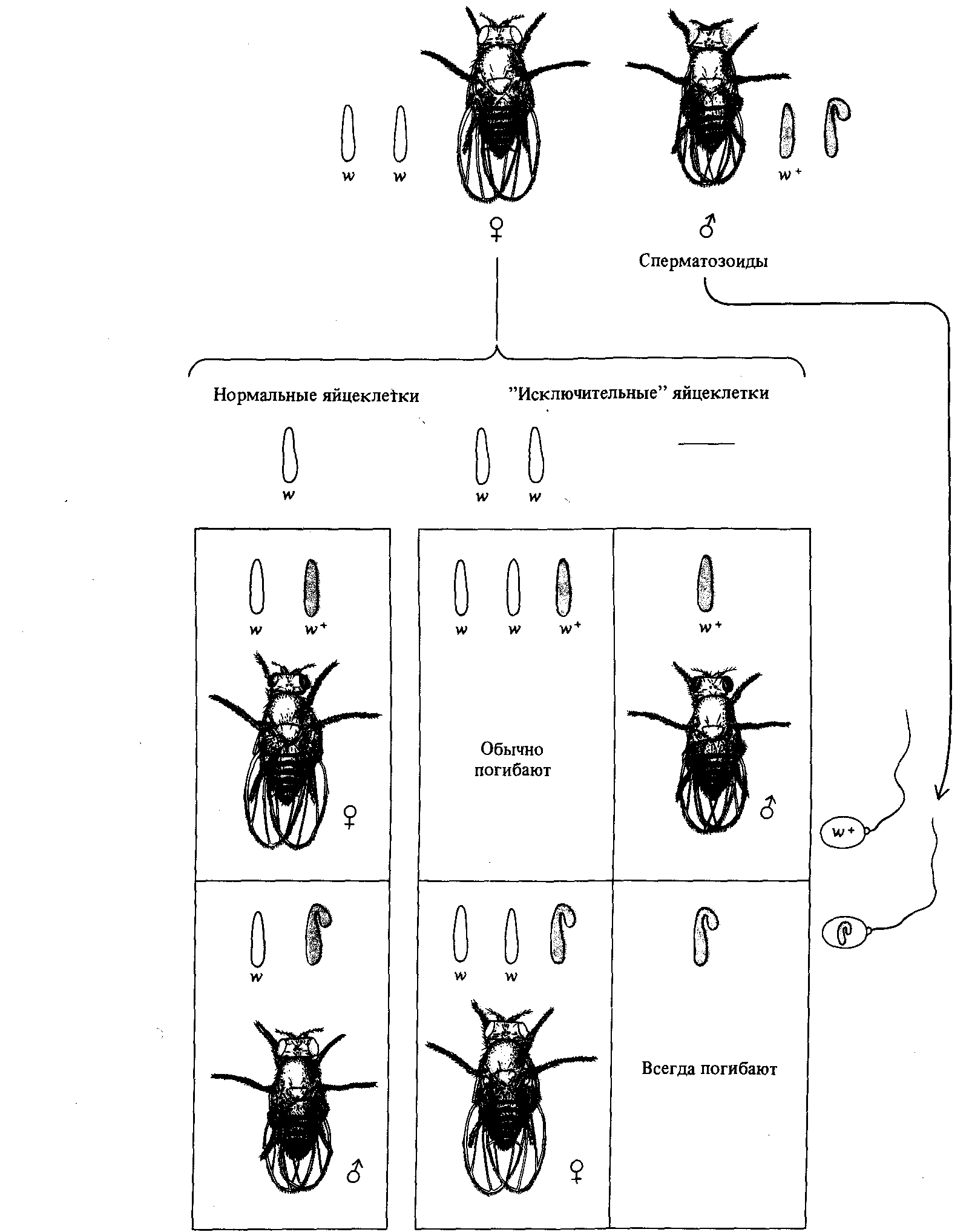
Как показал Морган, при скрещивании белоглазых самок дрозофилы с красноглазыми самцами дочери оказываются красноглазыми, а сыновья - белоглазыми (рис. 3.6). Однако и из этого правила бывают, оказывается, редкие исключения. Примерно у одной из двух тысяч мух в F1 от такого скрещивания цвет глаз оказывается противоположным: белым у самок и красным у самцов. Бриджес предположил, что появление редких мух возможно при нерасхождении Х-хромосом; другими словами, в тех случаях, когда Х-хромосомы не расходятся в мейозе к разным полюсам, а направляются вместе к одному из полюсов, в результате чего образуются яйцеклетки с двумя Х-хромосомами и яйцеклетки без Х-хромосом (рис. 3.7).
3. Хромосомные основы наследственности 71
|
|
|
Рис. 3.7. Первичное нерасхождение Х-хромосом у Drosophila melanogaster. В левой части рисунка для сравнения изображены результаты правильного расхождения. |
72 Организация и передача генетического материала
Если у белоглазой мухи образуется яйцеклетка с двумя Х-хромосомами и эта яйцеклетка оплодотворяется спермием, содержащим Y-xpoмосому, то образовавшаяся зигота имеет две Х-хромосомы и одну Υхромосому, причем обе Х-хромосомы содержат ген белых глаз. Бриджес предположил, что белоглазые самки в потомстве от скрещивания между красноглазыми самцами и белоглазыми самками развиваются именно из таких зигот. Когда же яйцеклетка, не содержащая ни одной Х-хромосомы, оплодотворяется спермием красноглазого самца, несущим Х-хромосому, то в зиготе оказывается одна Х-хромосома, несущая ген красных глаз, а Y-хромосомы нет вовсе. Бриджес предположил, что из таких зигот развиваются красноглазые самцы. Другими словами, гипотеза Бриджеса состояла в том, что появляющиеся с частотой 1 :2000 белоглазые самки получают по две Х-хромосомы от матери (и Y-хромосому от отца), а такие же редкие красноглазые самцы получают лишь одну отцовскую Х-хромосому (и, следовательно, вовсе лишены Y-хромосомы).
Гипотеза Бриджеса была умозрительной, но допускала экспериментальную проверку путем изучения хромосомных наборов мух, представляющих собой исключение из общего правила. Прямые наблюдения показали, что в клетках «исключительных» белоглазых самок действительно содержится по две Х-хромосомы и по одной Y-хромосоме, а клетки «исключительных» красноглазых самцов имеют по одной Xхромосоме, тогда как Y-хромосома у них отсутствует вовсе. Таким образом, было показано, что конкретный ген вне всяких разумных сомнений локализован в конкретной хромосоме.
Вторичное нерасхождение
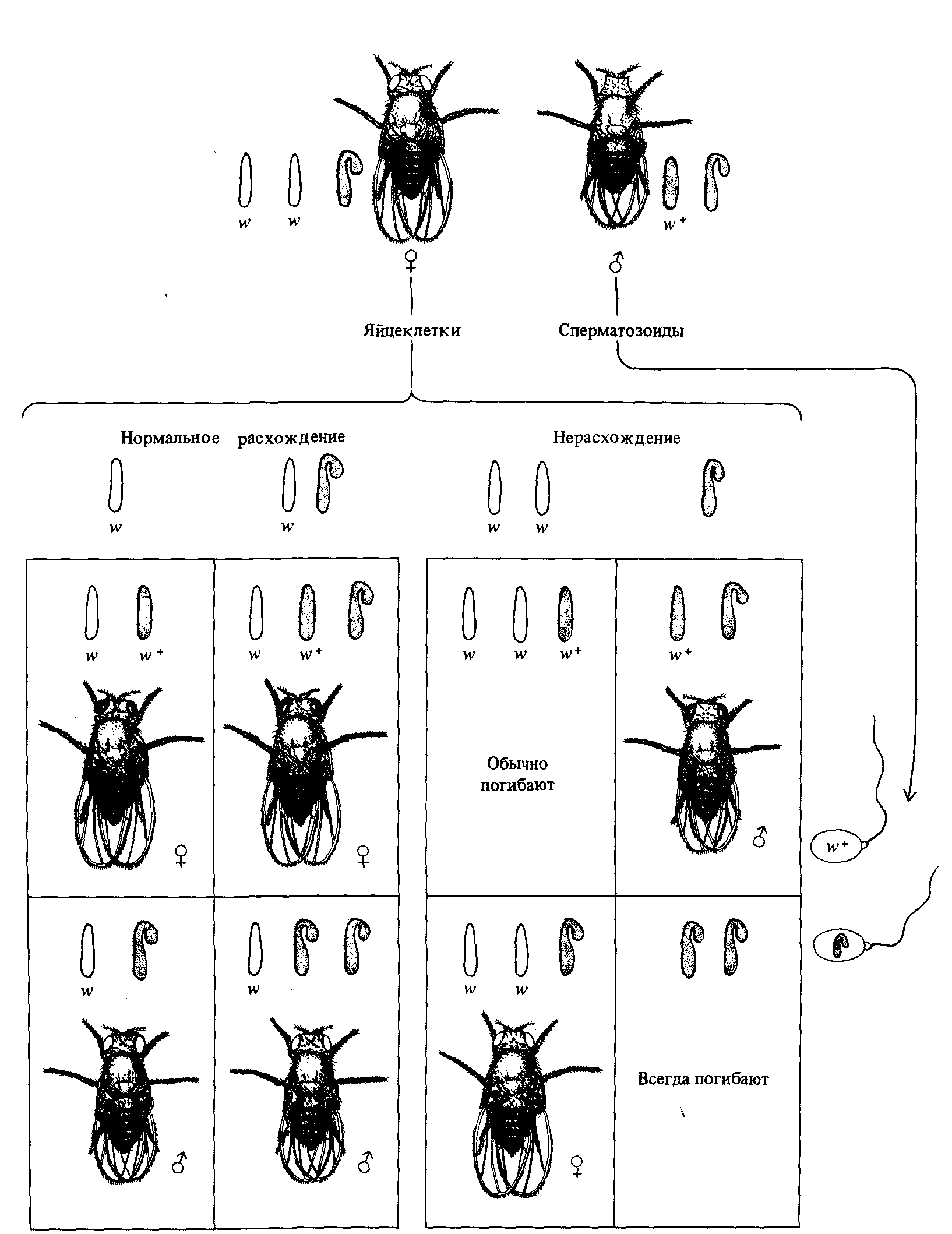
Самцы дрозофилы, лишенные Y-хромосомы, внешне нормальны, но стерильны. Самки с двумя Х-хромосомами и одной Y-хромосомой нормальны и плодовиты. Бриджес скрещивал таких самок (ΧΧΥ) с нормальными красноглазыми самцами (ΧΥ). Он обнаружил, что около 4% самок в потомстве от таких скрещиваний имеют белые глаза, а около 4% самцов - красные глаза; остальные 96% потомства составляли красноглазые самки и белоглазые самцы. Бриджес предположил, что эти 4% составляют самки и самцы, возникающие снова в результате нерасхождения Х-хромосом в мейозе у самок. Он назвал такое нерасхождение вторичным, поскольку оно происходит в потомстве самок, появившихся в результате первичного нерасхождения Х-хромосом (и потому обладающих двумя Х-хромосомами и одной Y-хромосомой) (рис. 3.8). Вторичное нерасхождение происходит с частотой около 1 :25, т. е. примерно в 100 раз чаще, чем первичное нерасхождение (1 :2000).
Нерасхождение может быть следствием физического сцепления Xхромосом, в таком случае нерасхождение имеет место в 100% случаев (это явление было открыто в 1922 году Лилиан Морган, женой Томаса Моргана) (рис. 3.9). Желтый цвет тела у Drosophila melanogaster определяется сцепленным с полом аллелем yellow (у), рецессивным по отношению к аллелю дикого типа (у + ). Цвет тела у дрозофил, гомозиготных по этому аллелю, много светлее нормального. Морган обнаружила существование желтых самок, которые при скрещивании с нормальными самцами давали в потомстве лишь желтых дочерей и нормальных сы-
|
|
|
Рис. 3.8. Вторичное нерасхождение Х-хромосом Drosophila melanogaster. Бриджес предположил, что мейоз у самок с двумя Х-хромосомами и одной Y-хромосомой происходит двояко: в первом случае Х-хромосомы расходятся к разным полюсам и к одному из них отходит также Y-хромосома (92%), во втором случае обе Х-хромосомы отходят к одному полюсу, а Y-хромосома -к другому; это происходит в 8% случаев, но только половина зигот, образовавшихся при участии таких гамет, способна к нормальному развитию, так что число соответствующих «исключительных» мух составляет около 4%. Свою гипотезу Бриджес подтвердил прямыми наблюдениями хромосомных препаратов различных типов мух. |
74 Организация и передача генетического материала
|
|
|
Рис. 3.9. Нерасхождение Х-хромосом вследствие их сцепленности. Х-хромосомы изображены цветными, Y-хромосомы - белыми. У желтых самок с такой двойной Х-хромосомой яйцеклетки бывают только двух типов, поскольку сцепление Х-хромосом сохраняется во время мейоза, и в результате они вместе отходят к одному из полюсов. При оплодотворении спермиями с Х-хромосомой эти яйцеклетки дают зиготы с тремя Х-хромосомами, которые обычно погибают, и зиготы с одной Х- и одной Y-хромосомой, как у обычных самцов. Когда те же два типа яйцеклеток оплодотворяются спермиями с Yхромосомой, то возникают зиготы с двумя сцепленными Х-хромосомами и одной Y-хромосомой и нежизнеспособные зиготы с двумя Y-хромосомами и не имеющие ни одной Х-хромосомы. |
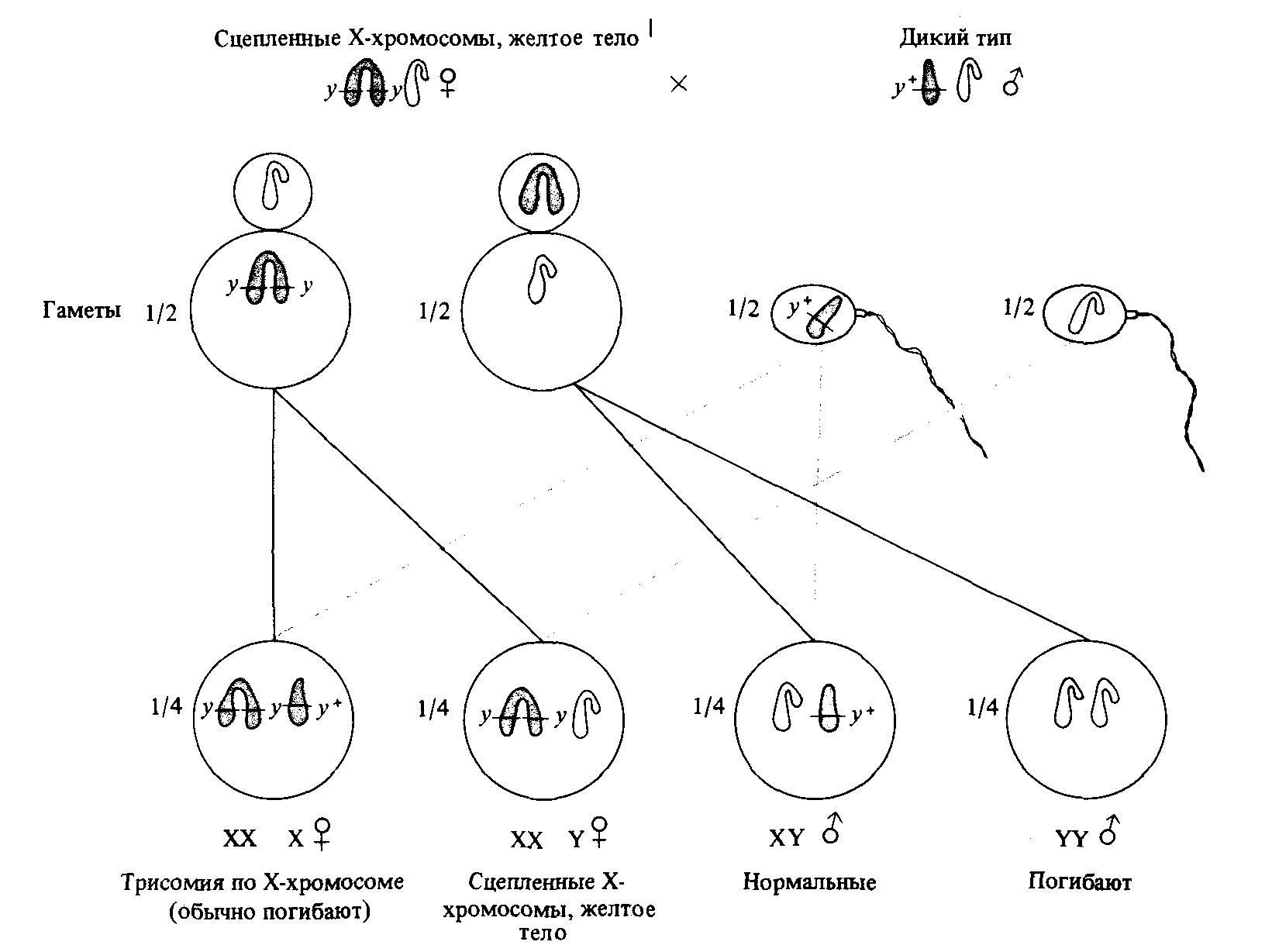
новей, а не нормальных дочерей и желтых сыновей, как подавляющее большинство. Желтые самки сохраняли эту особенность своих матерей: их потомство также составляли лишь желтые дочери и нормальные сыновья. Цитологический анализ показал, что клетки таких самок содержат по две Х-хромосомы, соединенных одной центромерой, и по одной Y-хромосоме.
Нерасхождение хромосом встречается не только у дрозофил, но и у других видов. У человека, например (см. гл. 21), нерасхождение в мейозе хромосом двадцать первой пары приводит к рождению детей, клетки которых содержат по три 21-х хромосомы. Это влечет за собой различные отклонения от нормального развития, называемые синдромом Дауна. Нерасхождение половых хромосом у человека приводит к нару-
3. Хромосомные основы наследственности75
шениям развития, называемым синдромом Клайнфельтера. Клетки таких людей содержат по две Х-хромосомы и по одной Y-хромосоме. Нерасхождения хромосом случаются также и в митозе.