

Генетические карты
Впервые порядок генов в хромосоме удалось установить Альфреду Стертеванту при выполнении им дипломной работы в «мушиной» лаборатории Томаса Ханта Моргана. В 1911 г. Морган и его студенты обнаружили существование множества сцепленных с полом мутаций у дрозофилы. Сцепленность с полом не оставляла сомнений в том, что соответствующие гены локализованы в Х-хромосоме. При проведении скрещиваний между линиями, содержащими различные мутации (см., например, рис. 5.6), Обнаружилось, что частоты рекомбинации между различными парами генов зависят от того, какая именно пара генов изучается, и почти постоянны для каждой пары. Частоты рекомбинаций, наблюдавшиеся между отдельными парами генов, представлены в табл. 5.1. Стертевант первым понял, что эти данные можно использовать для построения физической модели, или карты Х-хромосомы, на которой указывалось бы относительное расположение различных генов. Говоря более точно, Стертевант доказал на основании этих данных, что карта должна быть линейной, или одномерной, т. е. иметь форму, в точности совпадающую с нитевидной формой хромосом, наблюдаемой под микроскопом. Первая генетическая карта Х-хромосомы, построенная Стертевантом на основании данных, представленных в табл. 5.1, изображена на рис. 5.7.
Стертевант рассудил, что если гены расположены вдоль хромосомы, то чем дальше они расположены друг от друга, тем больше вероятность того, что кроссинговер произойдет между ними. Следовательно, частота рекомбинаций между удаленными генами должна быть больше, чем между соседними генами.
Проанализируем на основе этих соображений данные табл. 5.1. Частота рекомбинаций между мутациями yellow и white составляет 0,010, и, следовательно, они должны быть расположены много ближе друг к другу, чем yellow и vermilion, частота рекомбинаций между которыми составляет 0,322. Частота рекомбинаций между white и vermilion равна 0,300, следовательно, white должен быть ближе к vermilion, чем yellow. Аналогичное рассуждение показывает, что vermilion расположен рядом с miniature (частота рекомбинации 0,030) и что vermilion находится ближе к white, чем miniature (частоты рекомбинаций 0,300 и 0,327 соответственно). Следовательно, vermilion должен располагаться между white
|
Таблица 5.1. Частота рекомбинации (между некоторыми сцепленными с полом мутациями Drosophila melanogaster | |
|
Гены |
Частота рекомбинации |
|
Yellow (у) — white (w) Yellow (у) -vermilion (ν) Yellow (y) -miniature (m) Vermilion -miniature (m) White (w) -vermilion (v) White (w) -miniature (m) White (w) -rudimentary (r) Vermilion (v) -rudimentary (r) |
0,010 0,322 0,355 0,030 0,300 0,327 0,450 0,269 |
|
По Sturtevant A.H. (1913). J. Exp. Zool., 14, 43. | |
5. Геном эукариот135
|
|
|
Рис. 5.7. Первая генетическая карта Х-хромосомы D. melanogaster; на ней указано взаимное расположение генов yellow, white, vermilion, miniature и rudimentary. Ген yellow произвольно выбран точкой отсчета на генетической карте. |
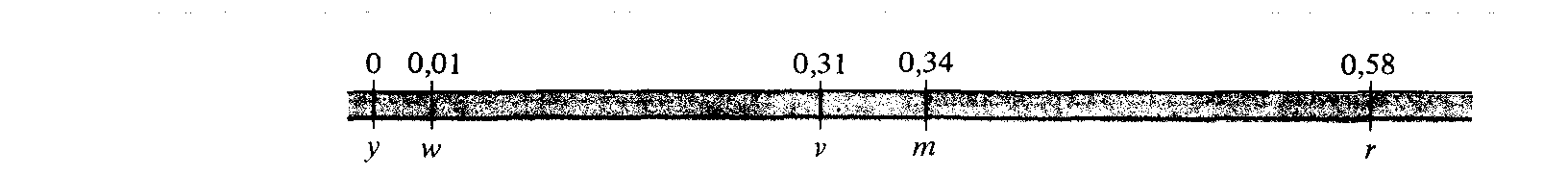
и miniature. Наконец, rudimentary почти не сцеплен с white, так как частота рекомбинаций между ними равна 0,45, т.е. близка к значению 0,50, отвечающему независимому расщеплению. Из того, что vermilion более тесно, чем rudimentary, сцеплен с white, следует, что он расположен между ними, и, значит, rudimentary лежит справа от white.
Взаимное расположение white и rudimentary показывает, что, как уже отмечалось ранее, два гена могут принадлежать одной хромосоме и тем не менее наследоваться независимо - для этого они лишь должны быть расположены достаточно далеко друг от друга. Например, ген, находящийся на некотором расстоянии справа от rudimentary, будет практически не сцеплен с white (т.е. частота рекомбинаций между ними будет неотличимой от 0,50), и тем не менее можно показать, что он принадлежит к той же хромосоме, что и white, поскольку оба они сцеплены с rudimentary. На рис. 5.7 расстояния между генами соответствуют частотам рекомбинации между ними. Частота рекомбинации, равная 1%, определяет единицу масштаба на генетической карте и называется морганидой или сантиморганидой (в честь Т. X. Моргана). Положение каждого гена на карте определяется его расстоянием от ближайшего из группы сцепленных генов. Произвольно принято, что ген yellow расположен на левом конце карты, т. е.находится в положении 0,0, и положение всех других генов оценивается по сумме частот рекомбинаций между ближайшими соседями. В результате получается, что расстояние между yellow и rudimentary на карте Х-хромосомы составляет 0,58 сМ.