

Тригибридные скрещивания
Мендель проверял закон независимого комбинирования на различных комбинациях пар признаков. Он подтвердил также этот закон, поставив опыт по скрещиванию растений, отличавшихся сразу по трем признакам. Такое скрещивание называется тригибридным.
Рассмотрим, например, скрещивание между двумя растениями гороха со следующими признаками:
|
Материнское растение: гладкие семена (RR) желтые семена (У У) пурпурные цветы (СС) |
Отцовское растение: морщинистые семена (rr) зеленые семена (уу) белые цветы (cc) |
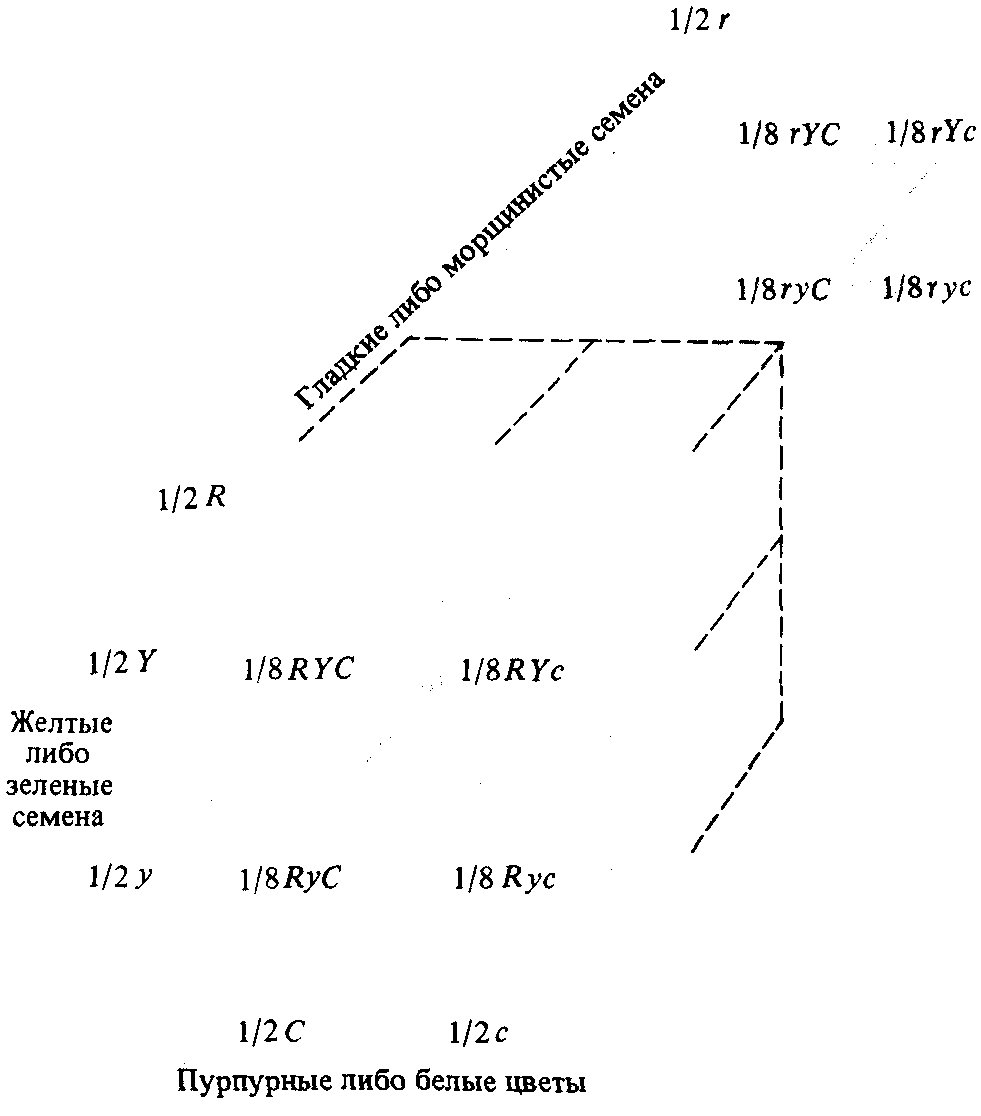
Материнское растение продуцирует гаметы типа RYC, отцовское — rус, следовательно, гибриды F1 будут тройными гетерозиготами или тригибридами, принадлежащими к генетическому типу RrYyCc. Вследствие доминантности семена у таких растений будут гладкими и желтыми, а цветы - пурпурными. Если все гены передаются независимо, то в тригибридном растении образуется восемь типов гамет, причем все с равной вероятностью (рис. 2.11).
|
Рис. 2.11. Гаметы, образующиеся у тригибридной особи. В отношении каждого гена вероятность одного из двух типов гамет равна 1/2. При одновременном рассмотрении всех трех генов возможны восемь типов гамет. Если все гены наследуются независимо, то вероятность каждого типа гамет составляет (1/2) ·(1/2)·(1/2) = 1/8. |
|
2. Менделевская генетика 51
|
|
|
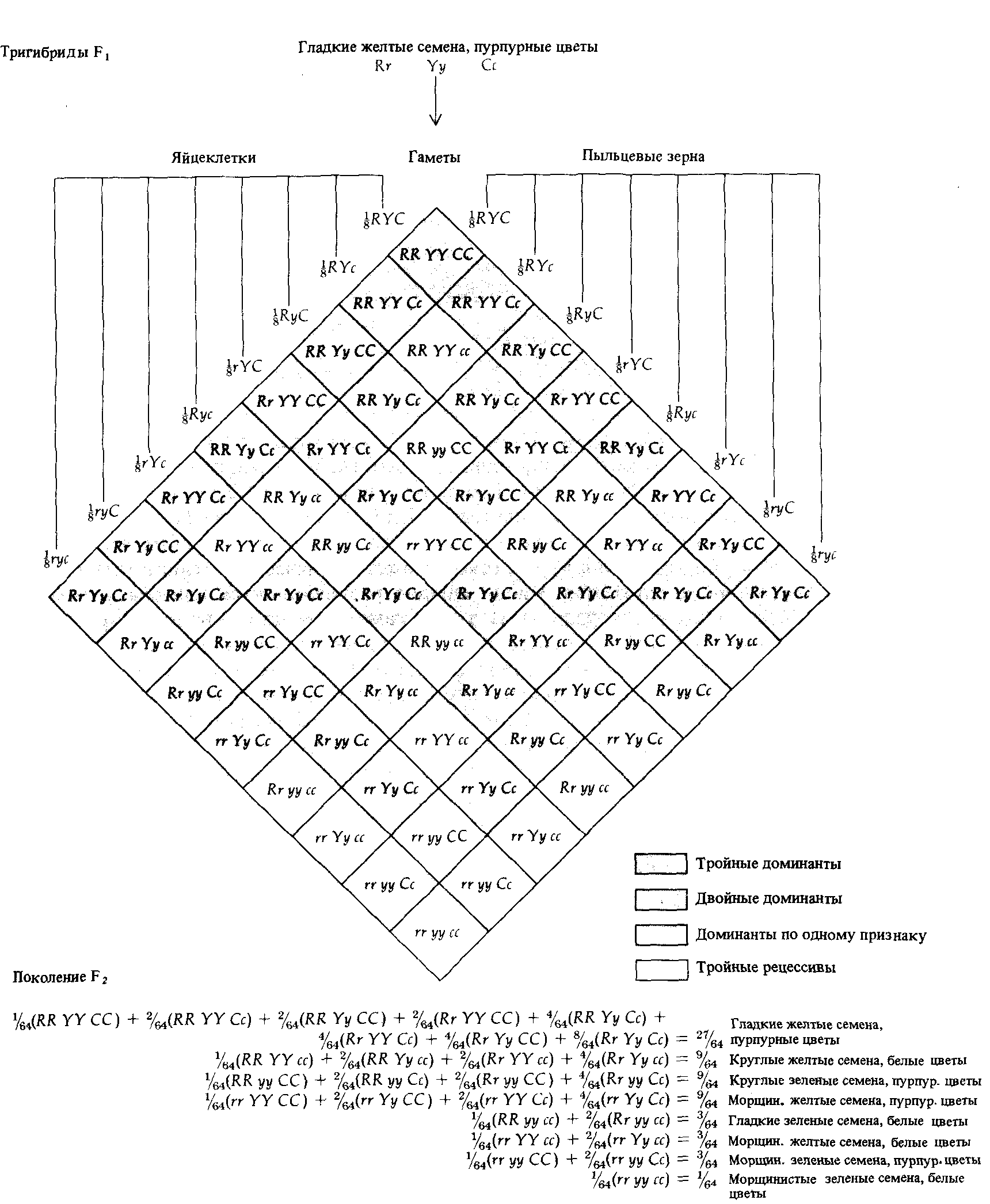
Рис. 2.12. Генотипы, возникающие в потомстве тригибридных особей при их самоопылении или перекрестном опылении. Существуют 64 комбинации восьми отцовских и восьми материнских гамет, но соответствуют они лишь 27 различным генотипам. В рассмотренном на схеме случае доминирования эти 27 генотипов соответствуют восьми различным фенотипам. В опытах Менделя тригибридные растения могли быть получены в результате скрещивания растений с гладкими желтыми семенами и пурпурными цветами и растений с морщинистыми зелеными семенами и белыми цветами. |
52 Организация и передача генетического материала
Случайное слияние гамет восьми типов от двух родителей приводит к возникновению 27 различных генетических классов (рис. 2.12). Вследствие доминантности этим 27 генетическим классам соответствуют всего лишь 8 типов внешне различающихся растений, представленных в следующем отношении (названия доминантных признаков выделены жирным шрифтом):
27 Гладкие желтые пурпурные
9 гладкие желтые белые
9 гладкие зеленые пурпурные
9 морщинистые желтые пурпурные
3 гладкие зеленые белые
3 морщинистые желтые белые
3 морщинистые зеленые пурпурные
1 морщинистые зеленые белые
Теперь мы можем сформулировать некоторые общие правила относительно потомства гибридов, полученных от скрещивания особей, отличающихся определенным числом генов (табл. 2.2). В общем случае каждый новый ген увеличивает число типов различных гамет вдвое, а число генетических классов (генотипов) втрое. Таким образом, особь, гетерозиготная по n парам генов, может произвести 2n типов гамет и 3n различных генотипов. Число внешне различающихся классов (фенотипов) равно числу различных типов гамет при наличии доминирования и числу различных генотипов в отсутствие доминирования.
Существует также несложная процедура, с помощью которой можно вычислить частоту данного генотипа в потомстве родителей, отличающихся определенным числом независимо наследуемых генов. Для этого надо подсчитать вероятности соответствующего генотипа для каждой пары генов отдельно, а затем перемножить. Допустим, мы хотим рассчитать ожидаемую частоту генотипа RryyCc в потомстве от скрещивания RrYycc х RrYyCc. Вероятность генотипа Rr в потомстве от скрещивания Rr х Rr равна 1/2; вероятность генотипа уу в потомстве от скрещивания Yy x Yy равна 1/4; наконец, вероятность генотипа Сс в потомстве от скрещивания cс x Cс равна также 1/2. Следовательно, вероятность генотипа RryyCc составляет (1/2)·(1/4)·(1/2) = 1/16·
|
Таблица 2.2. Число различных типов гамет в Ft и различных генотипов в F 2 при скрещивании особей, гомозиготных по двум различным аллелям определенного числа генов | |||
|
Число генов |
Число типов гамет |
Число генотипов |
Число фенотипов1) |
|
1 2 3 4 n |
2 4 8 16 2 n |
3 9 27 81 3n |
2 4 8 16 2 n |
|
1) При доминировании: в отсутствие доминирования число различных фенотипов совпадает с числом генотипов. | |||
2. Менделевская генетика 53