

Множественные аллели
Примеры наследования признаков, которые обсуждались до сих пор в этой главе и основывались на собственных опытах Менделя, касаютсядвухаллельных генов. Однако многие гены имеют несколько аллелей (множественный аллелизм), хотя каждый конкретный диплоидный орга-низм может быть носителем не более двух аллелей.
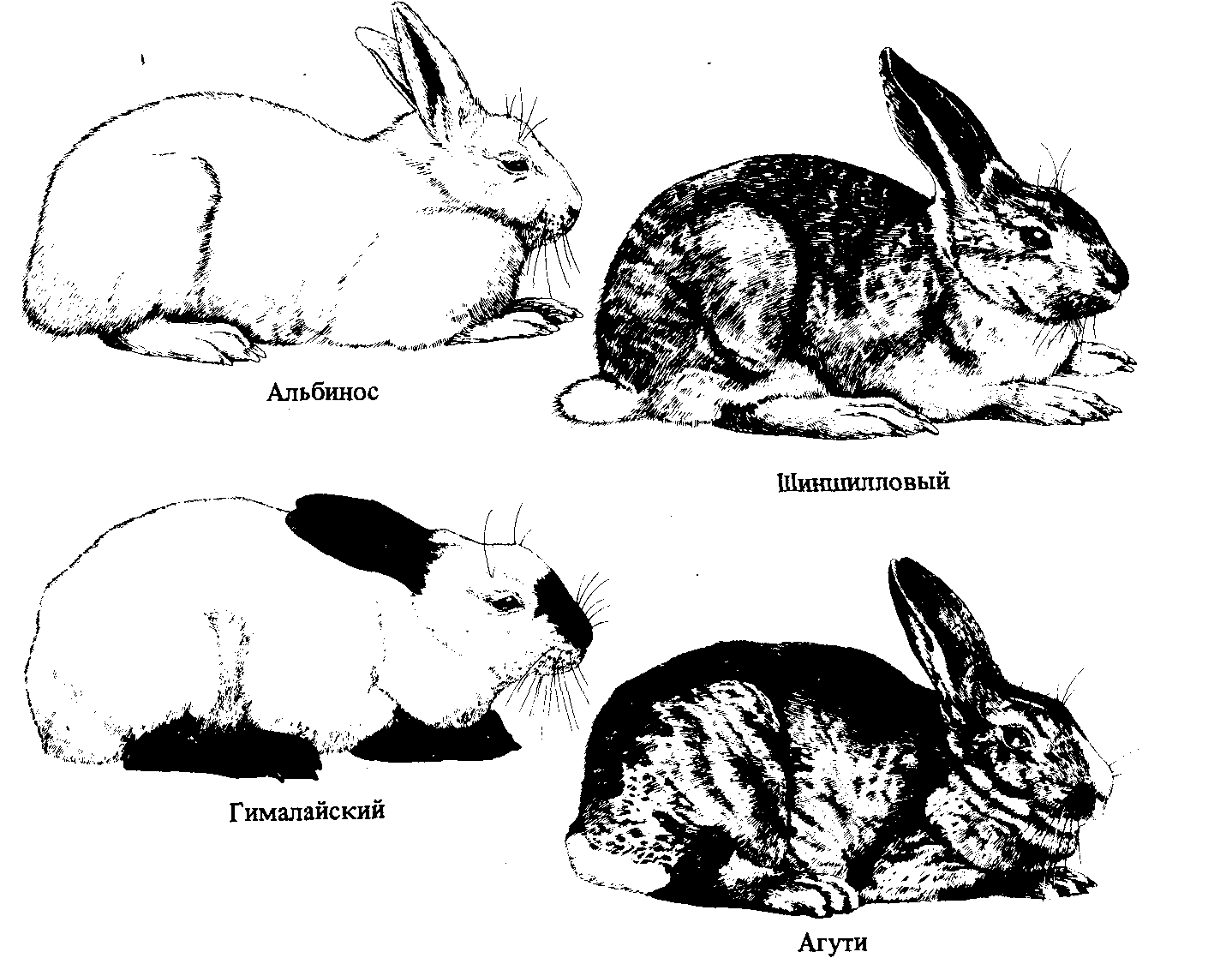
Известно большое число примеров множественного аллелизма; с не- которыми из них мы еще будем встречаться в этой книге. Одним из примеров может служить серия аллелей гена кролика, определяющего окраску меха; четыре из них приведены в табл. 2.3. Аллель дикого типа с+ доминантен по отношению к трем остальным: кролики, гомози- готные по с + или гетерозиготные по с + и любому другому аллелю, имеют обычную для этих животных серую (агути) окраску (или окраску
Таблица 2.3. Генетическое определение окраски меха у кроликов
|
Аллель |
Генотип |
Фенотип |
|
|
|
Дикий тип |
|
|
|
Шиншилла |
|
|
|
Светло-серый |
|
|
|
Гималайский |
|
|
|
Альбинос |
|
|
|
Рис. 2.13. Четыре фенотипа, возникающие при различных комбинациях алле- лей гена окраски меха кроликов. |
54 Организация и передача генетического материала
|
|
|
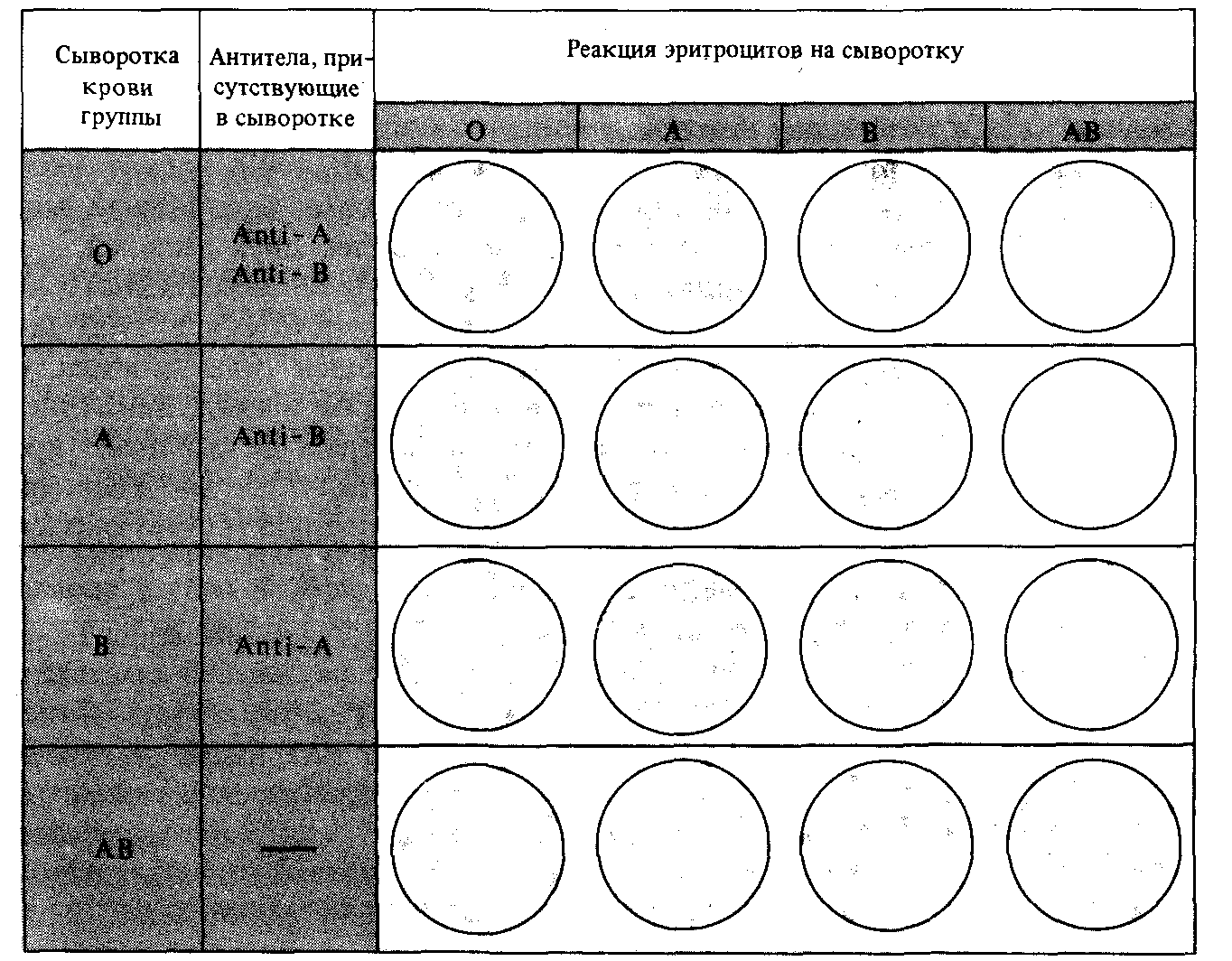
Рис. 2.14. Антигенные реакции, используемые при определении группы крови в системе АВО. В качестве тестера применяются сыворотки крови каждой из четырех групп. Наблюдается реакция, происходящая при смешении капли исследуемой крови с пробным раствором. Например, кровь человека с группой О не агглютинируется ни одним из четырех типов сыворотки, а кровь человека группы А агглютинируется сыворотками групп О и В. На агглютинацию указывает появление хлопьев. |
дикого типа). У особей, гомозиготных по аллелю ссh , мех по цвету напоминает мех шиншиллы и несколько светлее дикого типа. У гетерозигот по аллелям cch и сh или са мех светло-серый (промежуточный между шиншилловым и белым); аллель с проявляет, следовательно, неполное доминирование по отношению к аллелям сh и са. Гомозиготы сhсh и гетерозиготы сhсa - это кролики так называемого гималайского фенотипа, мех у них белый всюду, кроме лап, хвоста, ушей и кончика носа. Гомозиготы по аллелю са имеют типично альбиносный фенотип: белый мех и розовые глаза (рис. 2.13).
Другим примером множественного аллелизма может служить система групп крови АВО, открытая Карлом Ландштейнером (1868-1943) в 1900 году. Группы крови важно учитывать при подборе доноров для переливания крови, чтобы избежать слипания эритроцитов донора при их попадании в кровоток реципиента (рис. 2.14).
Существуют четыре группы крови системы АВО : О, А, В и АВ. Они определяются тремя аллелями одного гена : IА , IВ и i. Аллели IА и IВ доминантны по отношению к аллелю i, но кодоминантны по отношению друг к другу. При наличии трех аллелей возможно шесть генотипов; рецессивность i сводит число групп крови к четырем (табл. 2.4).
2. Менделевская генетика 55
Дополнение 2.1. Генетические обозначения
|
Система генетических обозначений развивалась без твердых заранее установленных правил, и это часто приводило к путанице. Генетики, работавшие с каким-либо определенным видом организмов, мало заботились о том, чтобы применяемые ими обозначения согласовывались с обозначениями, используемыми генетиками, работающими на других объектах. Ниже описываются принципы, которым мы будем следовать в этой книге при обозначении аллелей и генотипов. Если известны лишь два аллеля какого-то гена, то принято обозначать доминантный аллель курсивной прописной буквой латинского алфавита, а рецессивный-строчной. Например, три возможных диплоидных генотипа для пары аллелей А и α обозначаются как АА, Аа и аа. Однако в случае нескольких аллелей одного гена или когда известны независимые мутации гена, приводящие к одному мутантному фенотипу, обычно используются другие обозначения. Для обозначения гена или локуса используются курсивные строчные буквы (или группы букв), а аллели обозначаются индексом, помещаемым справа сверху. Например, буква с может обозначать ген окраски меха кролика. Нормальный аллель или аллель дикого типа (который часто бывает наиболее доминантным в серии множественных аллелей) обозначается символом с + , a другие аллели -символами сch, сh, сa и т.д. Часто обозначение с+ сокращают до знака « + ». При обозначении диплоидных генотипов мы обычно используем косую черту, чтобы показать, что каждый из двух аллелей находится в одной из двух гомологичных хромосом, например cch/c + или cch/ + . Удобство такого обозначения более очевидно при рассмотрении нескольких генов, расположенных в разных |
локусах одной хромосомы (см. гл. 5). Например, рецессивная мутация scarlet(st) в гомозиготном состоянии (st/st) определяет ярко-красный (алый) цвет глаз дрозофилы, тогда как доминантный аллель дикого типа st+ обусловливает темнокрасный цвет глаз. Рецессивная мутация ebony(e) в гомозиготном состоянии (е/е) определяет темный цвет тела, а соответствующий доминантный аллель дикого типа е+ -коричневато-желтый. Соответствующие два локуса расположены в одной хромосоме, и возможны два типа двойных гетерозигот, а именно st + е + /st е и st+ e/st e+. В первом случае говорят, что аллели st и е находятся в цис-положении (т. е. расположены в одной из гомологичных хромосом), а во втором случае -в транс-положении (расположены в различных гомологичных хромосомах). Изредка могут происходить доминантные мутации. Сочетания букв, обозначающие такие мутации, часто начинают прописной буквой. Например, мутация Ваr, определяющая полосковидную форму глаз у дрозофилы, доминантна по отношению к соответствующему аллелю дикого типа, обозначаемому символом В + . При обозначении аллелей, определяющих у человека группы крови системы АВО, используется необычная смешанная система обозначений: символы IА и IВ соответствуют кодоминантным аллелям, а буква г обозначает рецессивный аллель, в гомозиготном состоянии определяющий группу крови О. Описанные выше обозначения применяются как для гаплоидных, так и диплоидных генотипов эукариотических организмов. Обозначения, используемые в генетике бактерий, основаны на других правилах, которые будут изложены в главе 8. |
56Организация и передача генетического материала
|
Таблица 2.4. Группы крови системы АВО |
|
Таблица 2.5. Число различных генотипов при заданном числе аллелей | |||||
|
Аллель |
Генотип |
Фенотип (группа крови) |
|
Аллели |
Генотипы |
Гомозиготы |
Гетерозиготы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Количество различных генотипов при множественном аллелизме зависит от числа аллелей. Если аллель один, А, то и генотип один, АА. Если аллеля два, A1 и А2, то возможно три генотипа: два типа гомозигот а1а1 и А2А2 и гетерозиготы А1А2. При трех аллелях A1, A2 и А3 возможно шесть генотипов: три типа гомозигот А1А1, АгАг и А3А3 и три типа гетерозигот А1А2, А1А3 и А2А3. В общем случае при n аллелях возможно n(n + 1)/2 генотипов, из которых n-гомозиготы, а остальные n(п — 1)/2-гетерозиготы (табл. 2.5).