

Рестрикция днк и ферменты модификации
Практически все виды бактерий синтезируют по одному или по несколько типов специфических к определенной нуклеотидной последовательности эндонуклеаз, которые делают разрезы в двухцепочечной ДНК. Эти эндонуклеазы называются рестрицирующими ферментами (или рестриктазами), поскольку их основная функция состоит, по-видимому, в ограничении присутствия инородной ДНК в бактериальной клетке (рестрикциябуквально означает ограничение). ДНК клеток, синтезирующих ферменты рестрикции, защищена от их действия, потому что клетки синтезируюттакже модифицирующие ферменты, видоизменяющие структуру сайтов ДНК, узнаваемых ферментом рестрикции. Если клетка с действующей системой рестрикции и модификации инфицируется фагом с заранее не модифицированной ДНК, то вероятность того, что ДНК такого фага инициирует инфекцию, на несколько порядков меньше, чем для фага с модифицированной ДНК. Немодифицированная ДНК фрагментируется, число фрагментов зависит от числа сайтов узнавания в соответствующей молекуле ДНК, а затем фрагменты расщепляются экзонуклеазами. Изредка ферменты клетки-хозяина модифицируют фаговую ДНК до тогокак ее атакуют рестриктазы. В этом случае фаговая инфекция приводит к лизису клетки. Все потомки такого фага содержат тоже модифицированную ДНК и способны с высокой эффективностью заражать другие бактериальные клетки (с такой же системой рестрикции и модификации). Изучение закономерностей фаговой инфекции и привело к открытию системрестрикции и модификации ДНК и разработке методов получения чистых препаратов соответствующих ферментов.
9. Методы работы с ДНК267
|
Таблица 9.1. Последовательности, узнаваемые некоторыми рестрикционными эндонукле- азами |
|
|
|
1 Звездочками помечены основания, которые могут быть метилированы ферментами модификации. Малыми стрелками указаны разрезы, производимые рестриктазами типа II. Длинная вертикальная линия -ось симметрии. Pu -пурин, Ру- пиримидин. Размеры геномов: SV40-5224 н.п.; λ-48502κ.π.; аденовирус 2-38 200 н.п. |
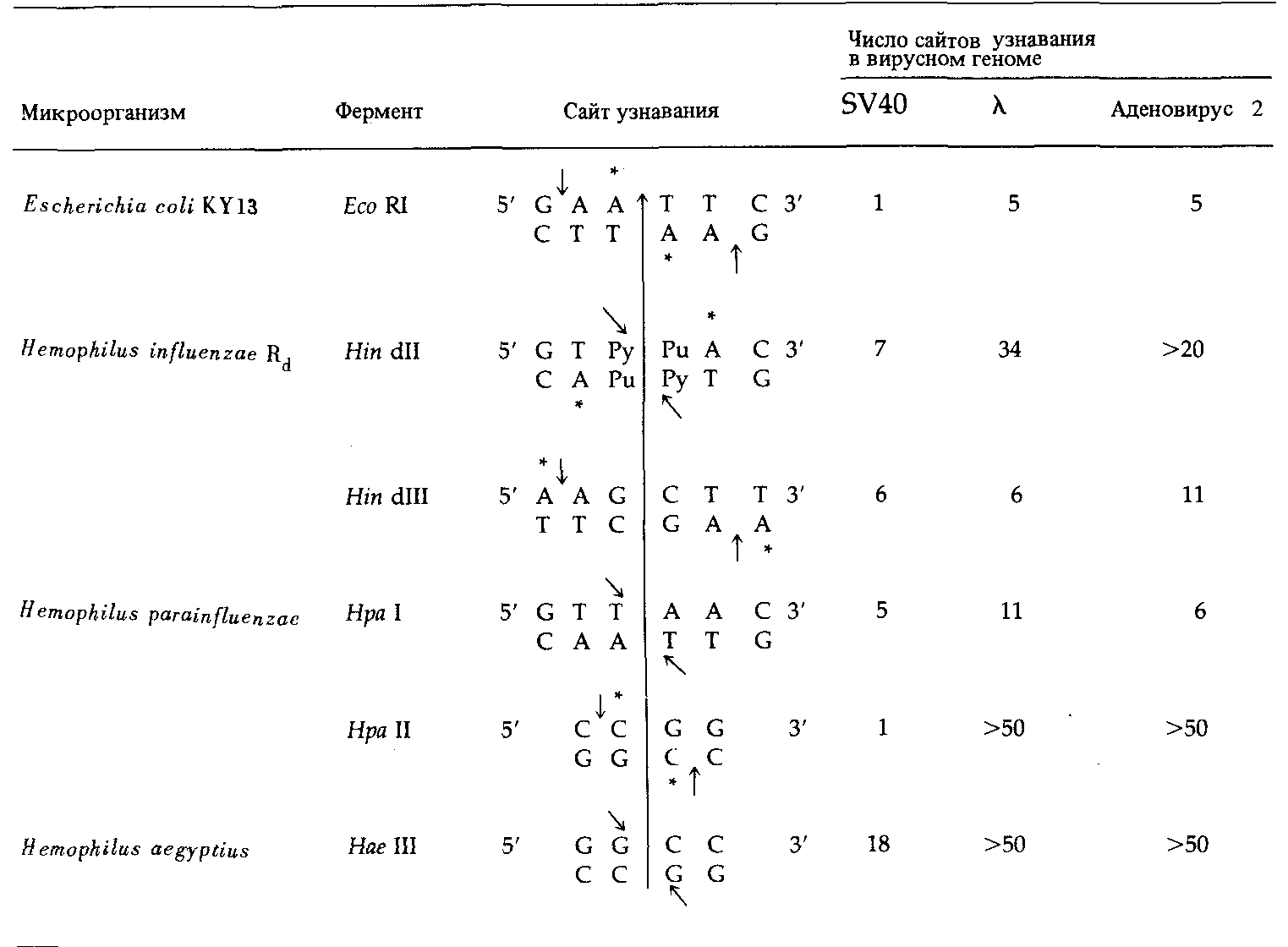
Известно три основных типа ферментов рестрикции. Рестрицирующие эндонуклеазы (рестриктазы) первого типа узнают определенную последовательность нуклеотидов и разрезают двухцепочечную молекулу ДНК неподалеку от этой последовательности, но само место разреза не строго специфично. Эндонуклеазы рестрикции второго типа узнают определенную последовательность и разрезают двойную спираль в определенной фиксированной точке внутри этой последовательности. Эндонуклеазы рестрикции третьего типа узнают нужную последовательность и разрезают двойную спираль, отступив определенное число нуклеотидных пар от ее конца. Мы в основном сосредоточимся на обсуждении свойств эндонуклеаз второго типа, поскольку именно они позволяют, во-первых, получать препараты ДНК, содержащие фрагменты с одинаковыми последовательностями нуклеотидов и, во-вторых, конструировать химерные молекулы ДНК, состоящие из фрагментов, взятых из разных геномов.
Рестриктазы второго типа узнают палиндромные последовательности-последовательности, обладающие центральной симметрией и считывающиеся одинаково в обе стороны от оси симметрии. Рестриктазы третьего типа, напротив, узнают асимметричные сайты. В табл. 9.1 представлены сайты узнавания для нескольких рестриктаз. Как указано стрел-
268Организация и передача генетического материала
|
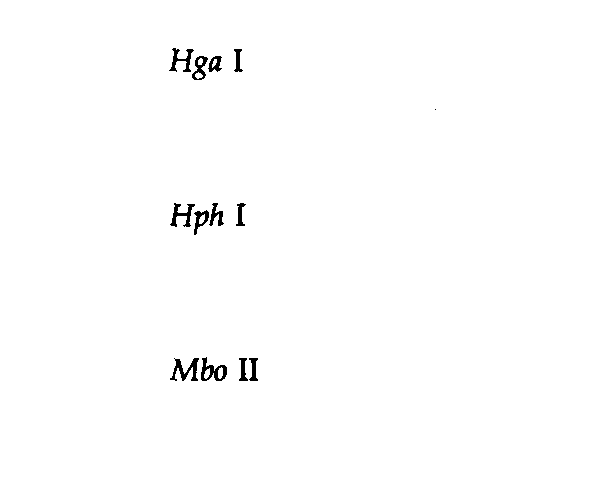
Таблица 9.2. Характеристика сайтов, узнаваемых рестриктазами | |
|
Фермент |
Сайт узнавания |
|
Тип Π (Ν = 7) |
|
|
Ek I |
|
|
Симметричный (Ν = 6) |
|
|
|
|
|
Вырожденный симметричный (Ν = 6) |
|
|
|
|
|
Симметричный (Ν = 5) |
|
|
|
|


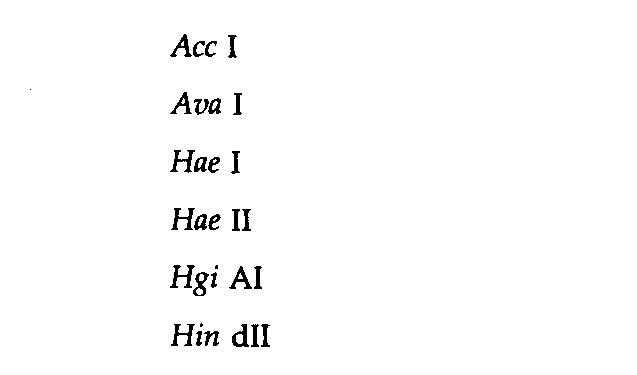

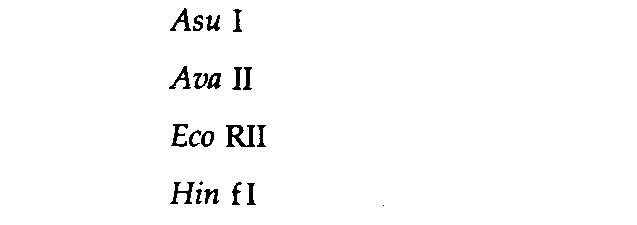
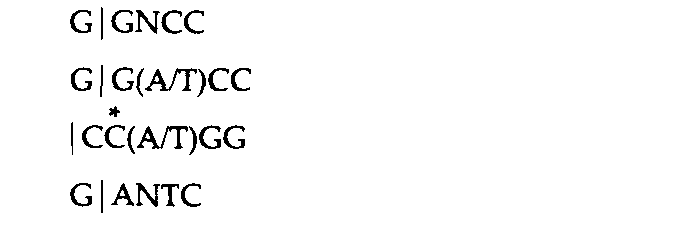
9. Методы работы с ДНК269
|
|
Продолжение табл. 9.2. |
|
Фермент |
Сайт узнавания |
|
Симметричный (N = 4) |
|
|
|
|
|
Симметричный метилированный (N = 4)++ |
|
|
|
|
|
Ассиметричный (N = 5) |
|
|
|
|
|
1 Для ферментов типа II последовательность представлена лишь в одной цепи; комплементарную последовательность можно достроить. Вертикальная линия обозначает положение, в котором разрываются фосфодиэфирные связи. Звездочки указывают основания, которые могут быть метилированы соответствующими ферментами модификации. 2 Специфический сайт узнавания Dpn Ι содержит метилированный аденин (mА), тогда как сайт Мbо I содержит неметилированное основание. Msp I узнает указанную последовательность независимо от метилированности внутреннего С; Ηρα II, напротив, узнает ту же последовательность лишь в том случае, если внутренний С не метилирован. | |

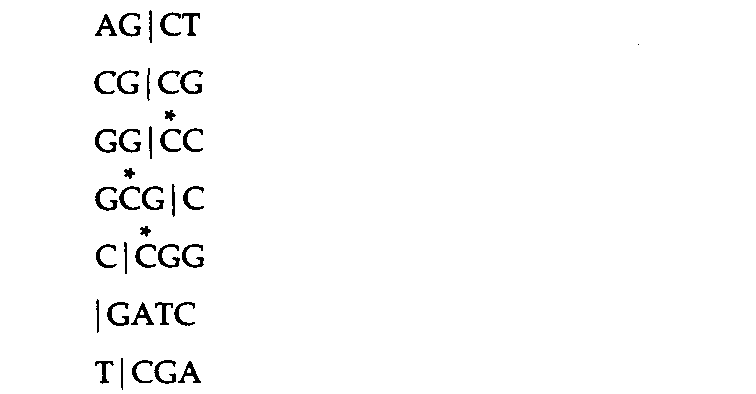
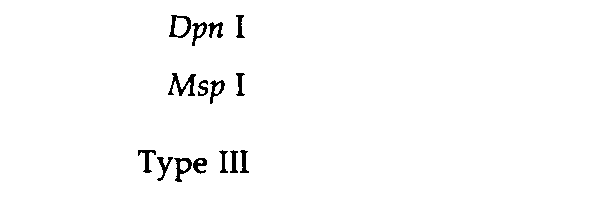

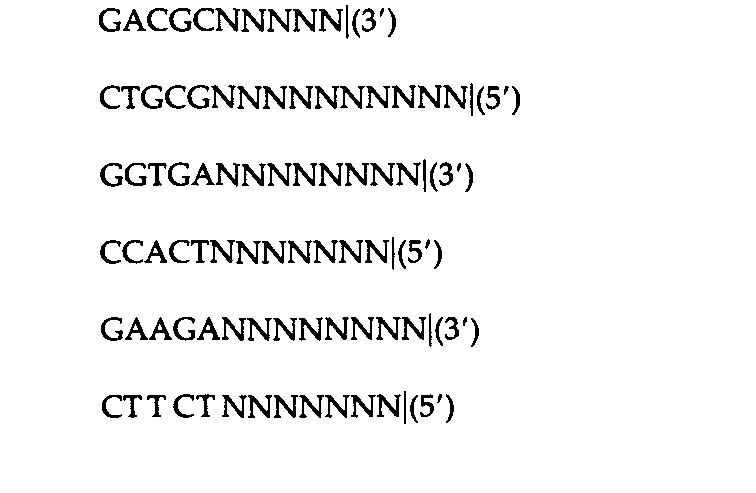
ками, точка разреза двойной спирали может совпадать с осью симметрии, а может быть сдвинута относительно нее. В последнем случае образуются комплементарные концы, и между соответствующими основаниями могут сперва устанавливаться водородные связи, а затем посредством ДНКлигазы происходить сшивка фрагментов с возникновением ковалентных связей между соседними нуклеотидами. Защита от повторного расщепления рестриктазой обеспечивается ферментами модификации, метилирую-
270 Организация и передача генетического материам
щими некоторые основания в сайте узнавания : соответствующие основания помечены звездочкой. Метилирование происходит после того, как соответствующий нуклеотид включается в ДНК в процессе репликации. Эти ферменты могут действовать либо на неметилированные, либо на полуметилированные сайты; при этом в процессе полуконсервативной репликации образуются полностью метилированные сайты.
Из различных видов и штаммов бактерий выделено и очищено более 175 различных рестриктаз, для которых известны сайты рестрикции. Выявлено более 80 различных типов сайтов, в которых происходит разрез двойной спирали ДНК. В таблице 9.2 приведена их классификация. Функция некоторых из этих ферментов почти наверняка состоит в защите клетки от присутствия чужеродной немодифицированной ДНК. Однако, в клетках некоторых видов бактерий эндонуклеазы рестрикции хотя и присутствуют, тем не менее, они, по-видимому, не ограничивают проникновение чужеродной ДНКinvivo. Вероятно, эти ферменты осуществляют какие-то иные функции. Как бы то ни было, рестриктазы независимо от их функций in vivo служат мощным инструментом структурного анализа геномов.