

МейозIi
К началу мейоза II хромосомы уже дуплицированы, и пары сестринских хроматид соединены общими центромерами. Однако каждая клетка содержит одинарный набор хромосом(N), а не двойной(2N), как в началемитоза или мейоза I. Профаза II часто проходит очень быстро. В метафазе II хромосомы прикрепляются центромерами к нитям веретена и располагаются в метафазной пластинке. К началу анафазы II каждая центромера делится (в первый и единственный раз в течение мейоза), и сестринские хроматиды таким образом становятся хромосомами, расходящимися затем к противоположным полюсам. Телофаза II завершается образованием ядерной мембраны вокруг каждого из двух гаплоидных ядер.
Мейоз I начинается в клетке, содержащей 2N удвоенных (дуплицированных) хромосом, и кончается образованием двух клеток (точнее, двух хромосомных комплексов, поскольку клеточное деление еще не совсем завершено), каждая из которых содержит по N дуплицированных хромосом. Мейоз II заканчивается образованием четырех клеток, каждая из
|
|
|
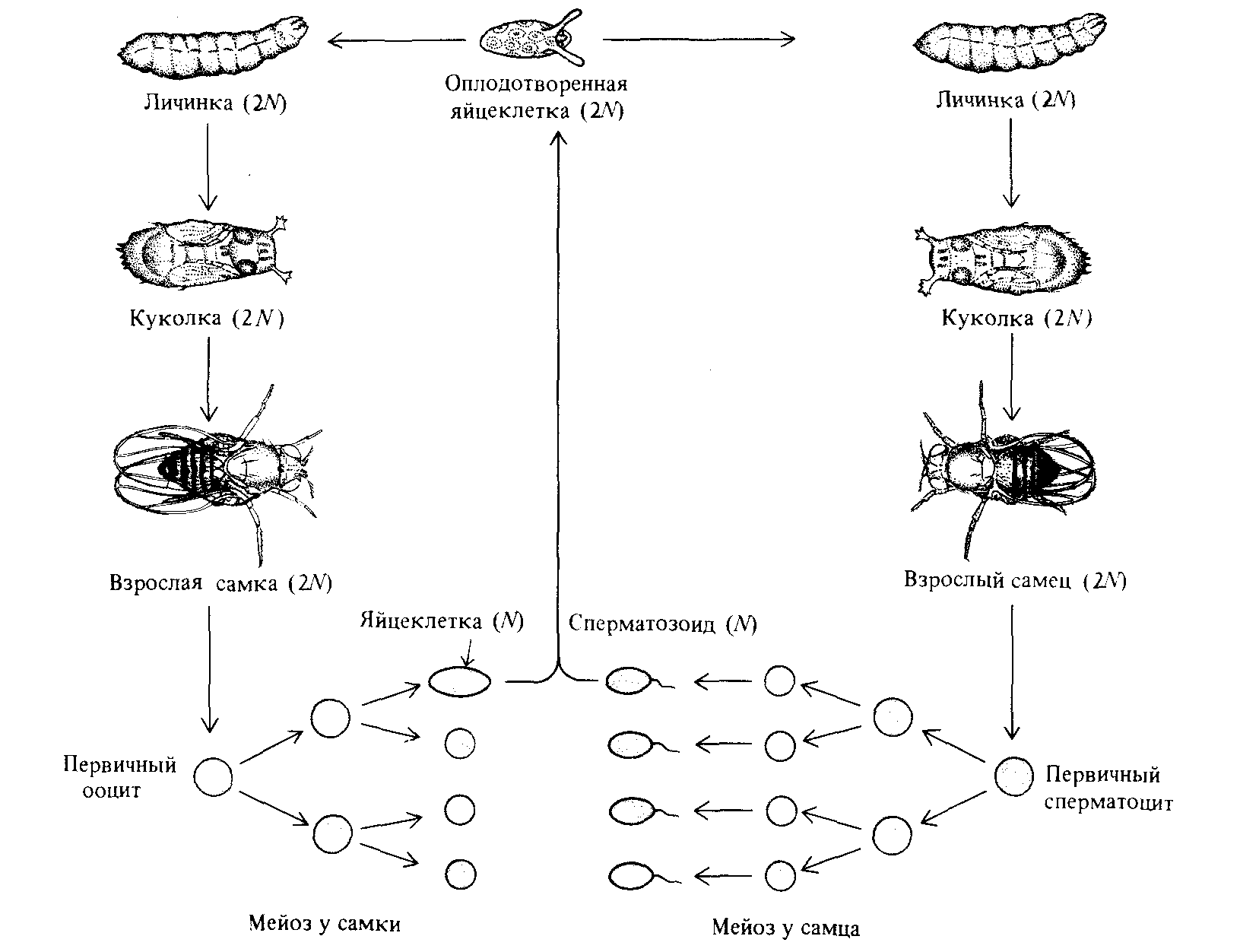
Рис. 1.16. Образование гамет у дрозофилы. Клетки зародышевой линии размножаются посредством митозов, а затем претерпевают мейоз и превращаются в ооциты (у самок) и в сперматоциты (у самцов). У самок лишь одна из четырех образующихся при мейозе клеток становится яйцеклеткой. Все четыре клетки, образующиеся в мейозе у самцов, называются сперматидами; в результате последующей дифференцировки они превращаются в сперматозоиды. |
32 Организация и передача генетического материала
|
|
|
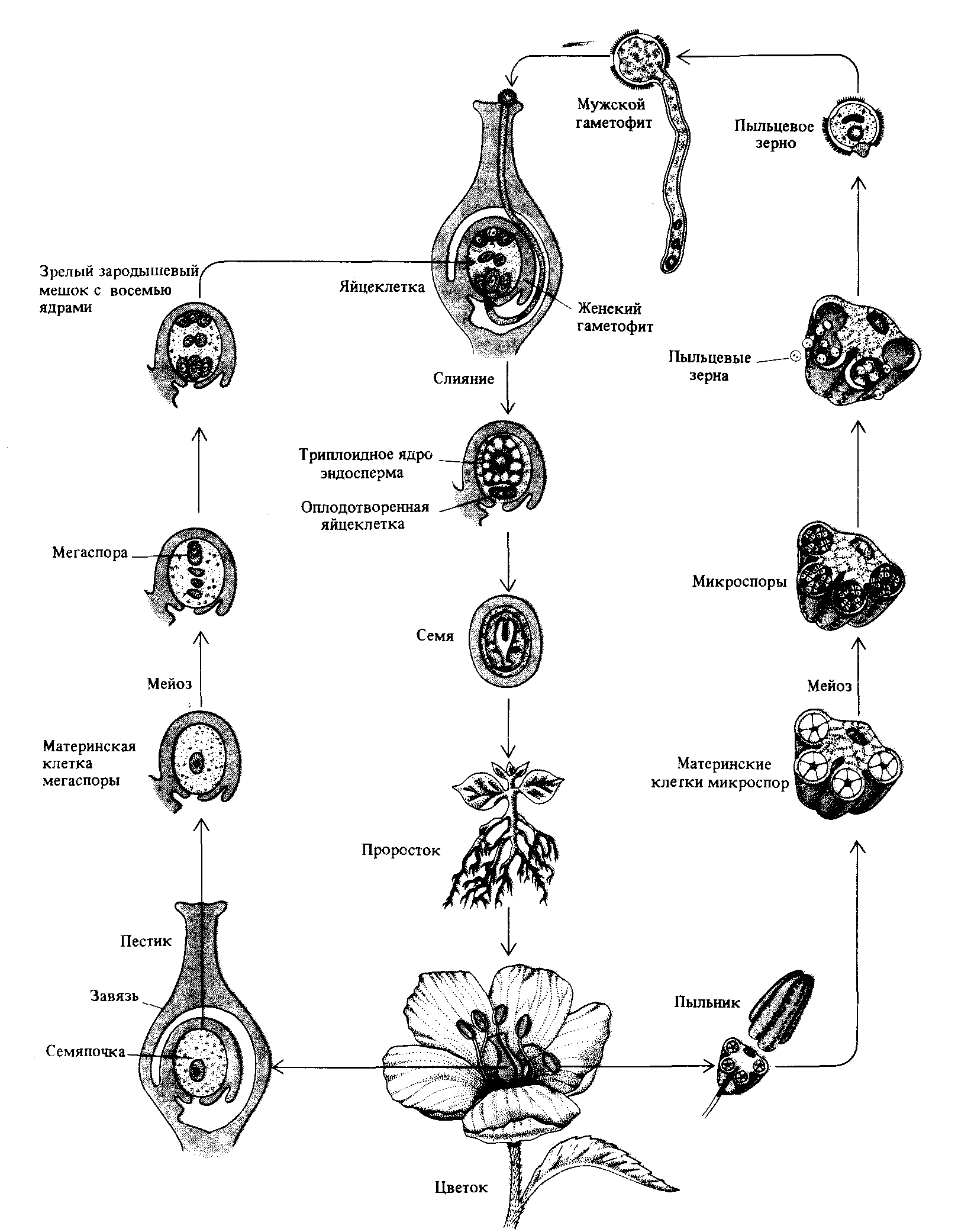
Рис. 1.17. Жизненный цикл и образование гамет у растения. У диплоида в результате мейоза происходит образование спор. Диплоидное растение называется спорофитом; гаплоидная фаза, включающая стадию созревания гамет, называется гаметофитом. Гаплоидная фаза может существовать в форме самостоятельного растения, независимого от диплоидной фазы. Например, у мхов гаметофиты представляют собой именно то, что мы называем мхом, тогда как спорофит представляет собой стебелек, живущий «паразитически» на гаметофите. |
1. Введение 33
которых содержит по N одинарных хромосом. Клетки, образуемые таким образом в мужских репродуктивных органах животных, носят название сперматозоидов. У самок, однако, лишь одна из четырех образующихся в мейозе клеток является яйцеклеткой ; остальные три клетки представляют собой полярные тельца, не способные выполнять функции гамет (рис. 1.16). У высших растений образующиеся в процессе мейоза мужские и женские половые клетки называются микроспорами и мегаспорами соответственно (рис. 1.17).
Значение мейоза
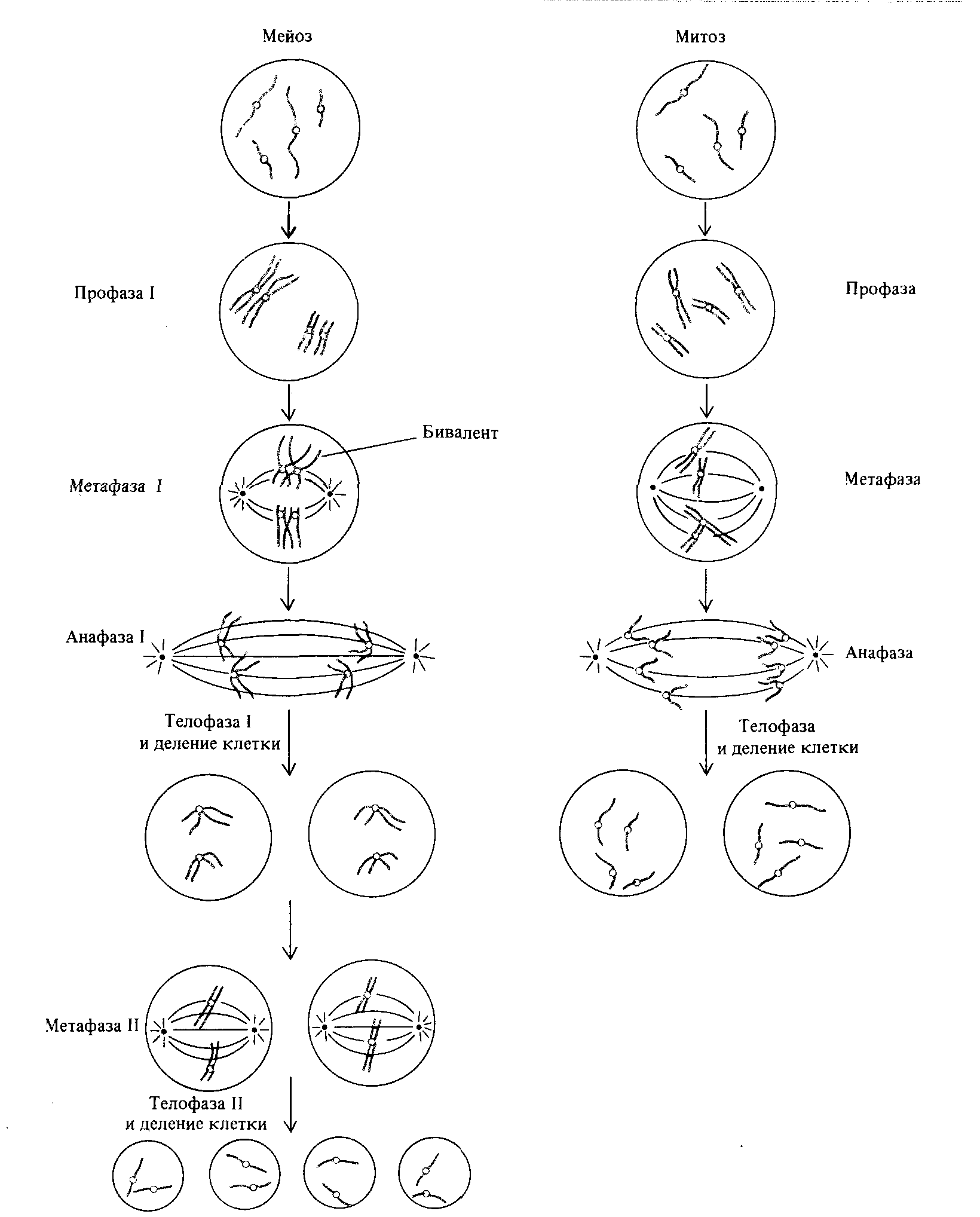
Митоз-это эквационное деление клетки, в результате которого хромосомные наборы дочерних клеток оказываются идентичными хромосомному набору родительской клетки. Иное дело мейоз: первое мейотическое деление является редукционным; второе-эквационным.
Мейоз I называется редукционным делением, поскольку число центромер и хромосом в клетках, образовавшихся в результате этого деления, вдвое меньше их числа в родительской клетке. Из каждой пары гомологичных хромосом родительской клетки одна из дочерних клеток получает отцовскую хромосому, а вторая-материнскую. Отцовская и материнская хромосомы могут содержать различную генетическую информацию; например, в отцовской хромосоме может содержаться информация «глаза карие, группа крови В», а в материнской - «глаза - голубые, группа крови О». Таким образом, клетки, возникшие в результате первого мейотического деления, генетически различны. Эти различия, однако, не всегда относятся к хромосоме в целом; каждый раз, когда несестринские хроматиды обмениваются участками, две хроматиды одной хромосомы становятся генетически различными (см. рис. 1.14).
С другой стороны, мейоз II - это эквационное деление, завершающееся делением центромер. Сестринские хроматиды - а, следовательно, и ядра, возникающие в результате мейоза II, - генетически идентичны (если не считать, как отмечалось в предыдущем абзаце, рекомбинантных участков). Существуют и некоторые другие различия между митозом и мейозом, которые можно видеть на рис. 1.18.
Генетическое значение мейоза можно суммировать следующим образом:
Мейоз обеспечивает постоянство числа хромосом у разных поколений организмов, размножающихся половым путем. Половое размножение включает стадию оплодотворения - слияния двух половых клеток или гамет. Если бы число хромосом в половых клетках было бы таким же, как и в соматических, то число хромосом удваивалось бы в каждом поколении.
В метафазе I каждая отцовская и материнская хромосома имеет равную вероятность оказаться по ту или другую сторону метафазной пластинки. Соответственно в каждой гамете могут оказаться как отцовские, так и материнские хромосомы. Если число хромосом значительно, то число возможных комбинаций сочетания отцовских и материнских хромосом в гамете очень велико, а вероятность того, что в определенную гамету попадут хромосомы только одного из родителей, очень мала. Рассмотрим, например, кариотип человека. В каждой нормальной клетке содержится 23 пары хромосом. Предположим, что первая отцовская хромосома оказалась по определенную сторону метафазной пла-
34 Организация и передача генетического материала
|
|
|
Рис. 1.18. Сравнение мейоза и митоза. В обоих случаях хромосомы однократно удваиваются, однако в мейозе происходит два клеточных деления, в результате чего число хромосом, приходящихся на одно ядро, уменьшается вдвое. Другое важное различие состоит в том, что в мейозе гомологичные хромосомы попарно объединяются, а в митозе - нет. |
1. Введение 35
|
|
|
Рис. 1.19. Четыре возможных типа взаимного расположения двух пар хромосом в метафазной пластинке. Две хромосомы, направляющиеся вместе к одному и тому же полюсу веретена, расположены в одном пятне; материнские хромосомы выделены более темным цветом, отцовские - более светлым. При одной паре хромосом число вариантов взаимного расположения равно двум; при двух парах - четырем (22=4), при η парах число вариантов составляет 2n. Заметим, что число различных вариантов результатов мейоза вдвое меньше числа взаимных расположений: результаты мейоза вариантов 1 и 2 одинаковы; то же относится и к вариантам 3 и 4. Вероятность того, что все отцовские (и соответственно все материнские хромосомы) направятся к одному полюсу веретена, равна (1/2)n – 1. Эта вероятность равна 1/2 для двух пар хромосом (п = 2), но быстро уменьшается с ростом числа хромосом. |
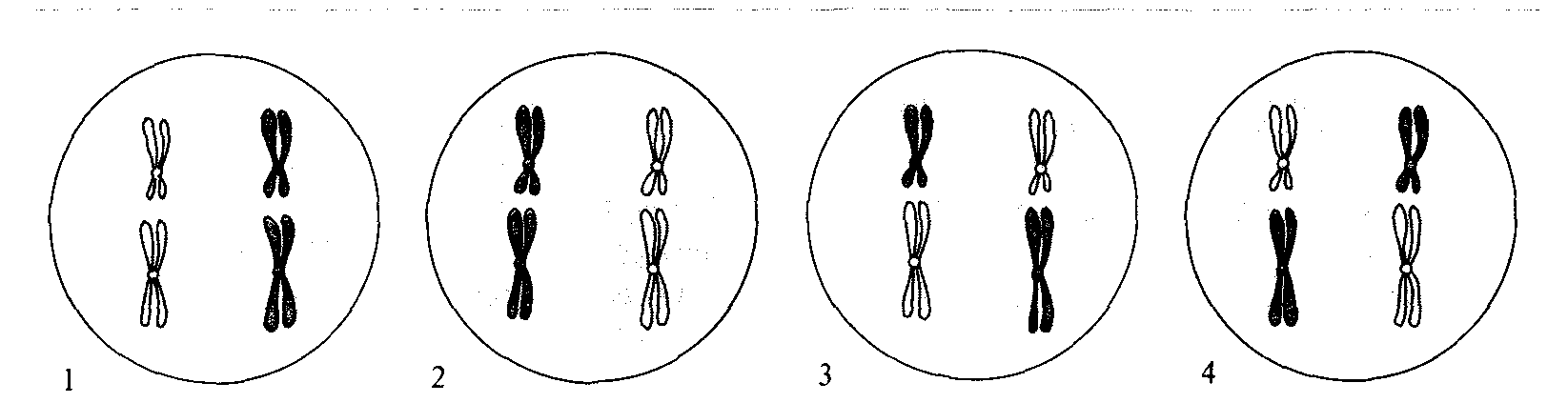
стинки. Вероятность того, что по ту же сторону пластинки окажется вторая отцовская хромосома, равна 1/2; то же самое справедливо для третьей, четвертой и всех остальных хромосом (рис. 1.19). Вероятность того, что все 23 отцовские хромосомы отойдут к одному полюсу, равна (1/2)22 = '/4 194 304, т.е. меньше одной четырехмиллионной.
3. Кроссинговер между несестринскими хроматидами еще больше перемешивает материнские и отцовские наследственные признаки в гаметах. В результате обмена участками между несестринскими хроматидами число различных типов гамет становится практически бесконечно большим. Напомним, что у человека в среднем на каждую хромосому приходится две-три хиазмы, а следовательно, два-три обмена участками хроматид. Границы этих участков от мейоза к мейозу варьируют, так что обмен генетическим материалом происходит каждый раз по-новому.