

Генетическое картированиеЕ. Coli
Метод прерванной конъюгации удобен при физическом картировании генов, довольно удаленных друг от друга, но не может использоваться при картировании маркеров, находящихся на близком расстоянии. Такие локусы картируют посредством рекомбинационного анализа, основанного на тех же принципах, которые были использованы при постановке трехфакторных скрещиваний (гл. 5 и 7).
Для рекомбинационного картирования требуется клетка реципиента с кольцевой ДНК хромосомы и ДНК донорной клетки. ДНК клетки-донора может вводиться в клетку реципиента различными способами: с помощью Hfr-хромосомы (при конъюгации), вместе с фагом-вектором (трансдукция), или путем прямой передачи ДНК из клетки донора в клетку-реципиент, как это описано в гл. 4 в отношении пневмококков (трансформация). Образующаяся при этом частично диплоидная клетка называется мерозиготой. Мерозиготы нестабильны, поскольку донорная ДНК представляет собой фрагмент целого репликона. Генетические
|
|
|
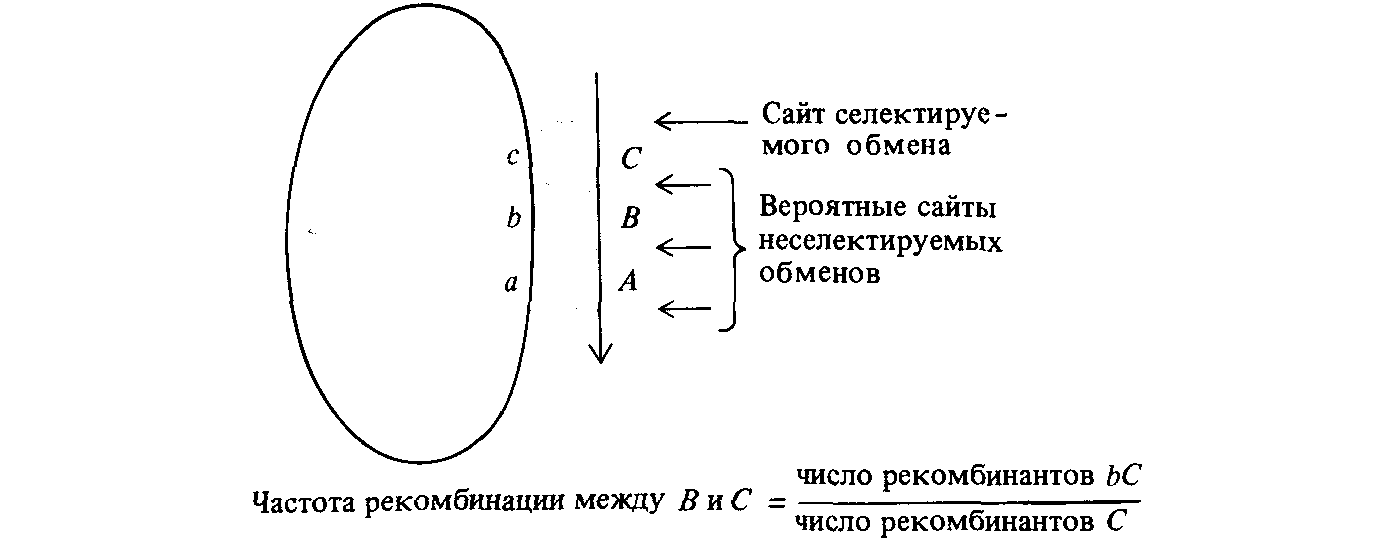
Рис. 8.14. Генетическое картирование посредством рекомбинационного анализа мерозигот, возникающих в результате конъюгации. Отбирается дистальный маркер С. Расстояние на генетической карте между В и С определяется как умноженное на 100 отношение числа рекомбинантов bС к числу рекомбинантов С. Поскольку в отличие от скрещивания между мейотическими организмами, реципрокные рекомбинанты в таком скрещивании не образуются, расстояния на генетической карте, определяемые таким образом, не аналогичны расстояниям, определяемым при скрещивании мейотических организмов. |
8. Бактериальный геном247
маркеры, содержащиеся в донорной ДНК, могут реплицироваться и сохраниться в потомстве, только если они попадают в репликон клеткиреципиента в результате рекомбинации. Как показано на рис. 8.14, для того, чтобы часть донорной ДНК встроилась в хромосому реципиента и при этом сохранилась кольцевая структура хромосомы, требуется два (или даже несколько) кроссинговеров.
Конъюгационное картирование
Когда производится картирование мерозигот типа F–, образовавшихся при конъюгации с клетками Hfr, полезно знать порядок попадания фланкирующих маркеров в клетку F–. Картирование выполняется посредством отбора по дистальному маркеру С (рис. 8.14). Эта процедура гарантирует участие в конъюгации всех интересующих нас маркеров. Частота появления неселективных маркеров у отобранных рекомбинантов используется для определения расстояния на карте от неселективного маркера до С, как это изображено на рис. 8.14. Например, тесно сцепленные мутации, влияющие на способность утилизировать лактозу в качестве источника углерода (lac Y, lac Z и lac I) в реципрокных скрещиваниях 6 и 7 передаются определенным штаммом Hfr после маркера pro + , но до маркера ade + .
Скрещивание 6: Hfr, Y–R Z+4 I+3 ade + strS x
F– , Y+R Z–4 I–3 ade– strR
Скрещивание 7: Hfr, Y+R Z–4 I–3 ade+ strS x
F–, Y–R Z+4 I+3 ade– strR.
Рекомбинанты ade + str* отбираются при посеве на агар с глюкозой и стрептомицином. Представленные в табл. 8.3 данные основаны на анализе генотипа более чем 10000 таких рекомбинантных колоний, от-
|
Таблица 8.3. Частоты неселективных маркеров среди рекомбинантов ade+strR, образуемых мерозиготами | |
|
Скрещивание 6 |
Скрещивание 7 |
|
| |
|
По Jacob F., Wollman E.L. 1961. Sexuality and the Genetics of Bacteria, Academic Press, New York, p. 229. | |

|
Скрещивание 6 |
|
|
|
Скрещивание 7 |
|
|
|
Рис. 8.15. Образование селективных и неселективных рекомбинантных генотипов в скрещиваниях 6 и 7 из таблицы 8.3. Все рекомбинанты содержат ген ade + (кроссинговер обозначен цветной стрелкой). Другие возможные обмены обозначены черными стрелками. Пунктирными стрелками обозначены кроссинговеры, положение которых по отношению к гену lacI не определено. |
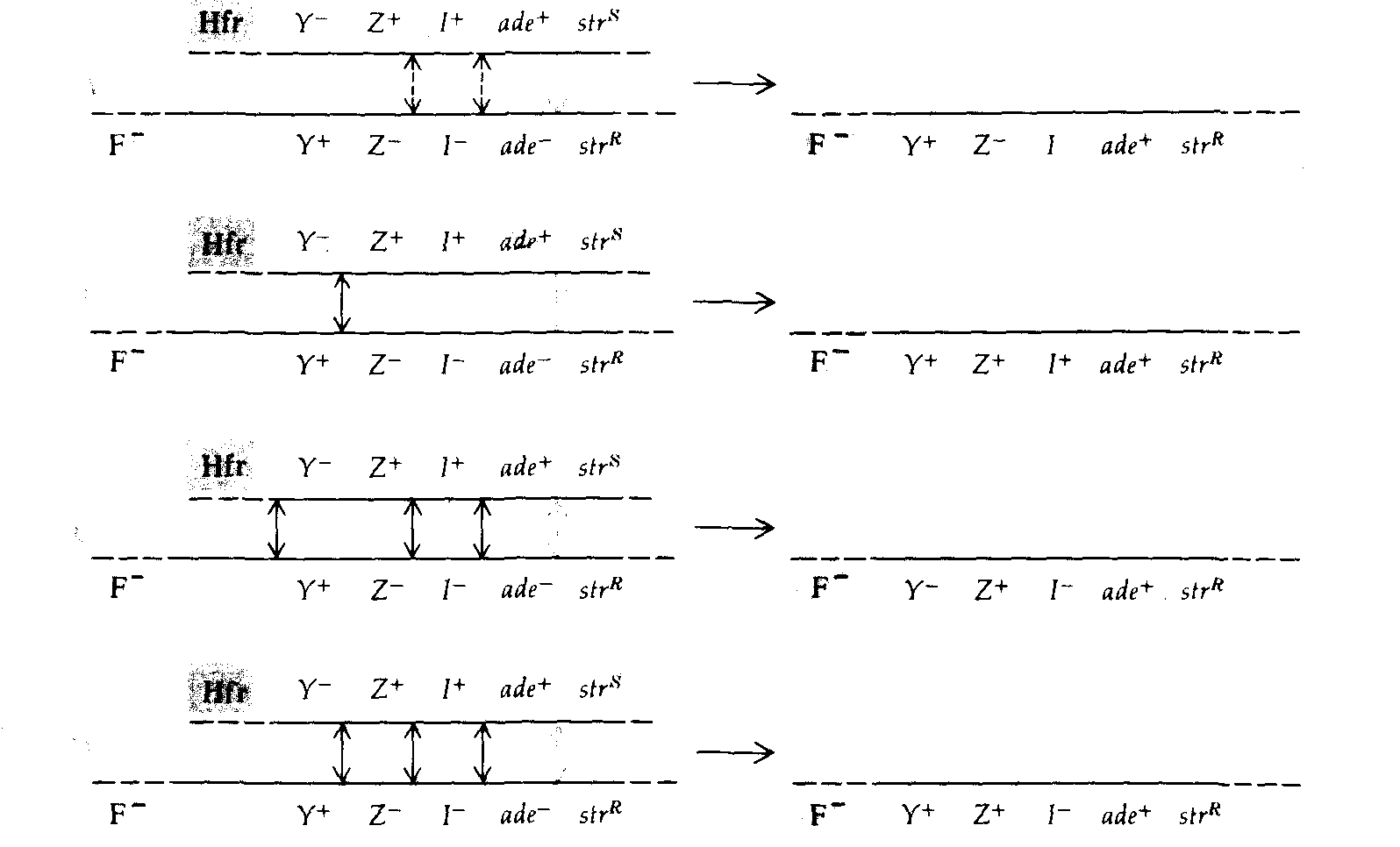
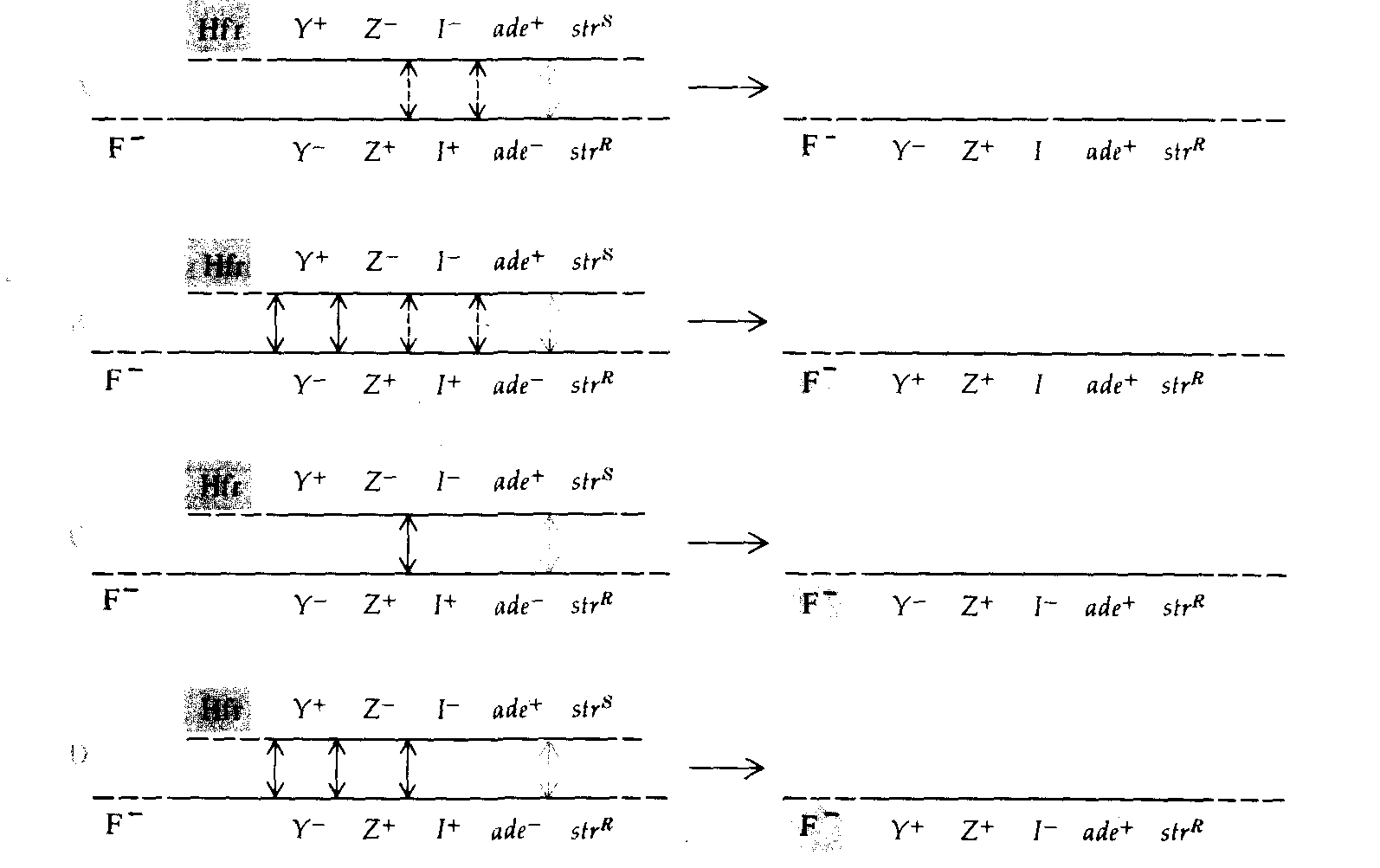
8. Бактериальный геном249
|
|
|
Рис. 8.16. Генетическая карта /ас-области Е. coli. |
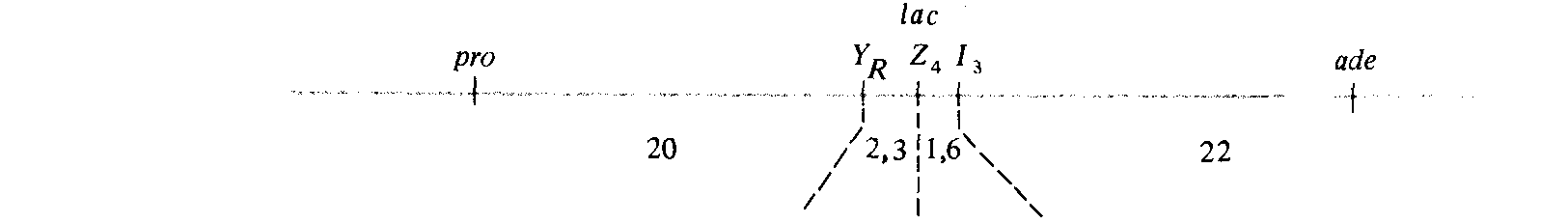
печатанных на чашки с питательной средой, позволяющей идентифицировать различные фенотипы Lac. Самый многочисленный класс рекомбинантов, полученных в этих скрещиваниях, позволяет определить расстояние на карте между ade и lacZ. В шестом скрещивании это расстояние оценивается в 22 единицы, в реципрокном седьмом скрещивании - в 26 единиц; такое соответствие можно считать удовлетворительным. На рис. 8.15 схематически изображены кроссинговеры, необходимые для возникновения рекомбинантов этого и других типов, представленных в табл. 8.3. Рекомбинанты классов В, С и D, полученные в скрещивании 6 (рис. 8.15), позволяют сделать вывод, что lad лежит между lac Z и ade, поскольку четырехкратный кроссинговер, в результате которого образуются классы С и D, должен происходить много реже двойного. Сравнение классов С и D с классом В показывает, что по аналогичной причине lac Ζ должен лежать между lacI и lac Y. Реципрокное скрещивание 7 подтверждает установленную в скрещивании 6 последовательность расположения генов. Полученные в этих скрещиваниях частоты четырехкратного кроссинговера свидетельствуют о существовании высокой отрицательной интерференции (см. гл. 14). На рис. 8.16 изображена рекомбинационная карта, построенная по данным, приведенным в табл. 8.3, и другим данным, полученным при использовании в качестве маркера аллелей локуса pro.
Сопоставление физической карты, полученной методом прерванной конъюгации, и генетической карты, построенной на основании данных о частоте рекомбинаций, показывает, что 1 минута продолжительности конъюгации соответствует 20 единицам генетической карты. Полная длина хромосомы Е. coli составляет 100 минут или 2000 единиц генетической карты. Одна единица карты соответствует примерно 1 600 н. п. Очевидно, что рекомбинационное картирование удобно лишь при малых расстояниях между исследуемыми локусами, поскольку маркеры, удаленные друг от друга более, чем на 3 минуты, расщепляются практически независимо, т.е. ведут себя как несцепленные гены.