

биотехнология кітап
.pdfтірі индивидуумдарда біртектес келеді. Мұның өзі қазір іс жүзінде қиындық келтірмейді деген сөз.
1971 жылы В.А. Струнников ген жəне хромосома деңгейіндегі генетикалық |
|
|||||||||||||||
инженерия əдістерін жібек құртында пайдаланды. |
|
|
|
|
|
|
|
|
|
|||||||
1972 жылы америка оқымыстысы П. Берг маймылдың онкогенді вирусының, |
|
|||||||||||||||
бактериофагтың жəне Е.СоІі бактериясы геномдарының |
түрлі бөліктерін |
біріктіру |
|
|||||||||||||
арқылы рекомбинантты ДНҚ алды. Міне, дəл осы зерттеу жұмысынан кейін, гендік |
|
|||||||||||||||
инженерия |
|
əдісі – |
биотехнология |
ғылымындағы болашағы |
үлкен |
сала |
ретінде |
|||||||||
жақсы қалыптаса бастады деп айтуға болады. Бірақ, мұндай құрастырмалы ДНҚ- |
|
|||||||||||||||
ның функционалдық активтілігі əрі қарай зерттелген жоқ еді, себебі мұндай жолмен |
|
|||||||||||||||
адам өміріне қауіпті бактериялар алынуы мүмкін деген |
күдік . |
тудыКейінірек |
|
|||||||||||||
зиянды зардаптары жоқ рекомбинантты ДНҚ алудың жаңа тəсілдері табылды. |
|
|
||||||||||||||
Қызметі жағынан активті, гибридті ДНҚ молекуласын ең бірінші болып1974 |
|
|||||||||||||||
жылы С. Коэн, |
Д. Хелинский, |
Г. |
Бойер |
алды. Олар |
|
құрамына |
|
əртүрлі |
|
|||||||
бактериялардың, құрбақа ооцитінің, теңіз кірпісі, тышқан митохондриясы ДНҚ-ның |
|
|||||||||||||||
фрагменттері |
енетін «химерлі» |
плазмидтерді |
құрастырып |
шығарды. Оларды |
|
|||||||||||
трансформация жолымен бактерия жасушасына енгізу |
|
арқылы, онда |
жоғары |
|
||||||||||||
сатыдағы организмдер ДНҚ-на тəн транкрипция құбылысын жүргізуге мүмкіндік |
|
|||||||||||||||
туылды. Кейінірек активті қызмет атқаратын рекомбинантты ДНҚ-ын Кеңес Одағы |
|
|||||||||||||||
ғалымдары С.И. Алиханян мен А. А. Баст құрастырды. |
|
|
|
|
|
|
|
|
||||||||
Жасуша деңгейінде жүргізілетін жұмыстардың ішінде сомалық жасушаларды |
|
|||||||||||||||
гибридизациялау əдісі кеңінен қолданылады. 1960 жылы Ж. Барский жануарлардың |
|
|||||||||||||||
сомалық жасушалары бір-бірімен қосылысуға, яғни екі жасушадағы генетикалық |
|
|||||||||||||||
информация бірігуге қабілетті екендігін көрсетті. |
|
|
|
|
|
|
|
|
|
|||||||
К. Гржешик (1973 ж.) көп |
жыл |
бойы |
зертханада |
жүргізген тəжірибелерінің |
|
|||||||||||
нəтижесінде, денелік (сомалық) жасушалардың |
түр |
ішіндік |
жəне |
|
түр |
аралық |
||||||||||
гибридтерін алды. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Денелік жасушаларды гибридтеу өсімдіктерге де жүргізілді. Темекі, сəбіз, |
|
|||||||||||||||
қызанақ |
сияқты |
өсімдіктердің |
|
жеке |
протопластарынан |
жасуша |
қабық |
|||||||||
регенерацияланып, каллус түзілгеннен кейін тұтас өсімдік қалыптаса алатындығы |
|
|||||||||||||||
анықталды. |
|
Осы |
тұрғыдан |
|
көптеген |
|
зерттеушілер |
|
|
протопластарды |
|
|||||
генетикалық |
информацияны |
қабылдайтын |
жақсы |
реципиент |
|
жəне |
де |
|||||||||
жасушаларды будандастыру үшін қолайлы материал деп біледі.
1953 ж. АҚШ ғалымы Джеймс Уотсон мен ағылшын физигі Фрэнсис Крик ДНҚ-ның қрылымын зерттей келе, тұңғыш рет оның шиыршықталып келген екі спираль тəрізді пішіндегі нобайын жасаған. Бұлар өз жұмыстарында Морис Уилкинс жəне Розалинда Франклиннің(Ұлыбритания) ДНҚ-ның рентгенді құрылысына сараптама жасау жəне Эрвин Чаргаффтың(АҚШ) ДНҚ-ның нуклеотидті құрамы
жөніндегі |
мəліметтеріне сүйенді. Мұндағы нуклеин |
қышқылдарындағы негізгі |
|
«құрылыс |
материалдары» мононуклеотидтер |
болып |
табылатын. Əрбір |
мононуклеотид бір азотты негізден (пуринді немесе пиримидинді), пентозадан, жəне
фосфат қалдығынан құралған. Оған |
моншақ |
тəрізді«орналастырылған» 4 типті |
||
нуклеотидтер (аденин-А, тимин-Т, |
гуанин-Г |
жəне |
цитозин)-Цқатал |
түрде |
белгіленген тəртіп бойынша орналастырылған жəне бір-бірімен жұп құрады. Аденин тек тиминмен жұп құрса, гуанин тек цитозинмен серіктеседі. Мысалы,
10
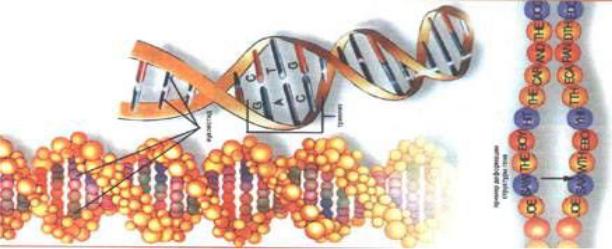
жылқылардың геномында шамамен осындай жұптың3 биллионы бар. Осындай ретпен орналасқан спиралдың əрбір жұбы өзінің«серіктесі» жайлы толық мəлімет береді. Осылайша, бір жұптың арқасында оған серіктес екінші жұпты жасауғ болады. ДНҚ-ның молекулярлы моделін келесі суреттен көре аламыз (сурет-2).
2-сурет. ДНҚ-ның молекулярлы моделі.
Генетикалық инженерияның жасушалық жəне ағзалық деңгейлері бір-бірімен
тығыз байланысты. Жануарлар жасушаларымен |
жүргізген тəжірибелерде бір |
|
жасушаның ядросын |
екіншісімен алмастыруға, екі немесе бірнеше эмбрионды |
|
қосып біріктіруге, оларды бірнеше бөлікке бөлшектеуге болатындығы анықталды. |
||
Мысалы, генотиптері |
əртүрлі ұлпалардың |
жасушаларын біріктіру жолым |
тышқанның аллофенді даралары алынған. Ол үшін дамудың бастапқы бластомерлі кезеңіндегі эмбриондар алынады жəне оларды бөліп алған соң проназа ферментінің көмегімен жеке-жеке бластомерлерге бөлшектейді. Содан соң екі немесе бірнеше ұрықтың бластомерлерін қосып, соның негізінде тұтас бір комплексті эмбрион жасалынады. Сөйтіп, ақ жəне қара тышқандар бластомерлерінің бірігуі нəтижесінде
– аллофенді ұрық дамиды. Гаструла стадиясында ұрық пробиркадан алынып, аналық тышқанға жіберіледі. Одан туатын ұрпақ екі түрлі ата-ананың фенотиптерін біріктіретін аллофенді болып шығады. Аллофенді ұрпақтарды үш, төрт немесе одан да көп ата-анадан да алуға болады.
Денелік жасушаларды біріктірумен қатар, бір жасушадан бөлініп алынған жеке хромосоманы, басқа жасушаға енгізудің де жолдары табылған. Сондай жеке бөлініп
алынған |
хромосома гендерінің, басқа, яғни бөтен жасушаға барғанда да ө |
|||
қызметтерін атқаратындығы анықталған (Мак-Брайд, Озер, 1971). |
|
|||
|
2. Гендік инженерия əдістерінің мазмұны |
|
||
Ген |
инженериясының |
теориялық |
негізіне– генетикалық |
кодтың |
универсалдылығы жатады. Бір ғана кодтың (триплеттің) барлық тірі ағзалардың – адам, жануар, өсімдік, бактериялардың ақуыз молекулаларының құрамына енетін амин қышқылдарын бақылай алатындығына байланысты, ДНҚ молекуласының кезкелген бөлігін басқа бөтен жасушаға апарып салу, яғни молекулалық деңгейде
11
гибридтеу, теориялық тұрғыдан алғанда мүмкін болып есептеледі. Генді бөтен жасушаға апарып салу(трансгеноз) жəне геннің өзін ДНҚ құрамынан бөліп алу немесе қолдан синтездеу, əрине, өте күрделі жұмыстар. Ал ағзаға (организмге) сырттан енгізілген геннің, жаңа генетикалық аппарат құрамына қосылып қызмет атқаруы, одан да күрделірек. Дегенмен бұл салада қол жеткен табыстар баршылық. Мысалы, жасанды ортада балапан ұрығының жасушасы өсірілген. Белгілі-бір уақытта оған жаңа синтезделген ДНҚ жіпшелерінің құрамына бромдезоксиуридин косылған. Осылайша таңбаланған жаңа синтезделген ДНҚ-ын бұрынғы ескі ДНҚ-нан оңай ажыратуға болады. Сонымен қатар, осындай ортаға тышқан жасушасынан бөлініп алынған тритиймен(3Н) таңбаланған ДНҚ қосылған. Содан біраз уақытта, жасуша көбейгеннен кейін, оның құрамындағы генетикалық материалды алып қарағанда, тышқанның ДНҚ-ы мен балапан ДНҚ-ның араласып кеткендігі байқалған.
Гендік инженерия əдістерімен рекомбинантты ДНҚ құрамына енетін жекелеген гендерді мынандай жолдармен дайындауға болады:
1)табиғи ортадан (жасуша, организм) тікелей бөліп алу;
2)химиялық жолмен синтездеу арқылы алу;
3)белгілі-бір генге сəйкес келетін рРНҚ-ның көшірмесін алу.
Бірінші əдіс ген инженериясы дамуының бастапқы кезеңінде ке қолданылды. Бұл əдіс бойынша түрлі организмдердің жасушаларынан бөліні
алынған |
тұтас |
ДНҚ |
моекулалары |
рестриктаза |
ферменттерінің |
кө |
|
бөлшектелініп, реципиент |
жасушаларға |
жіберіледі |
жəне |
олардан |
г |
||
молекулалардан тұратын иондар алынады. Бүл əдіс осы күнге дейін өз мəнін жойған |
|
||||||
жоқ, мысалы, қазір гендердің банкін жасау үшін пайдаланылуда.
Генді химиялық жолмен қолдан синтездеу тұңғыш рет 1969 жылы Г Корананың зертханасында жүзеге асырылды. Г Корана өзінің қызметтестерімен бірге ашыту бактериясының көмегімен аланиңді рРНҚ генін синтездеді. Ол ген бар жоғы77 нуклеотидтен тұрған жəне реттеуші механизмі жоқ болғандықтан, активті қызмет атқара алмаған. Кейінінен олар функционалдық жағынан активті200 нуклеотидтен
тұратын |
тирозинді рРНҚ генін синтездеді. Қазіргі кезде қолдан |
синтезделген |
гендердің |
ішіндегі ең ұзыны– адамның өсу гормонының гені |
жəне 584ол |
нуклеотидтен тұрады.
Генді жасанды жолмен алудың үшінші əдісі– кері транскрипция арқылы жүретін ферментативтік синтезге негізделген. Бұл ең алғаш онкогенді вирус РНҚның репликациясын зерттеу барысында анықталды. Сонда кері транскриптаза ферментінің көмегімен ИРНК матрицасының негізінде ген синтезделген.
Осындай жолмен адам |
мен |
жануарлардың жəне |
құстардың глобинде, |
||
жұмыртқа |
ақуызын, |
сиырдың |
көз |
айнасының ақуызын |
жəне. коделейтін.б |
(кодтайтын) |
гендер |
ашылды. Бүл əдіс |
адам интерферонының генін бөліп алып, |
||
бактерия жасушасына жіберу үшін де қолданылды. Интерферон — вирустық инфекциямен жəне басқа аурулармен, соның ішінде қатерлі ісікпен күресу үшін қолданылатын тиімді емдік дəрмек. Интерферон жануарлар мен адам жасушасынан да жасалады. Ю. А. Овчинников пен В. Г. Дебасов адам интерферонын синтездейтін микроорганизмдерді алды. Алдымен олар адам интерферонын синтездеуге қабілетті рекомбинантты ДНҚ-ын құрастырып алды да, содан соң оны бактерия жасушасына
12
жіберді. Ондай бактериялар 1 литр суспензияға шаққанда5 мг интерферон синтездей алады. Бүл 1 литр қанның құрамындағыдан 5000 есе көп.
Гендік инженерияның əдістерін қолдану үшін, хроммен жақсы өңделген қожайын – вектор қажет.
Вектор – белгілі организмде дербес репликацияланушылық қабілеті , бар сонымен бірге оған бөгде ДНҚ-ның енуіне кедергі келтірмейтін, ДНҚ-ның шағын молекуласы. Мұндай қабілет бактериофагтар мен плазмидтерде байқалады.
Екінші шарт – микроорганизмдерге векторлық жəне рекомбинантты молекулаларды енгізудің тиімді тəсілі болуы қажет. Өнеркəсіпте гендік инженерия əдістерін, микробтық синтез көмегімен медицинада қолданылатын адамдар ақуызын жəне ветеринарияда қажетті ауылшаруашылық малдарының ақуыздарын өндіруге мүмкіндік туды. Мысалы, белгілі-бір аурулармен зақымданғандардың организіміне
тиісті |
ақуыздарды – |
интерферон, |
полипептидтік |
гормондарды, иммунно- |
модуляторларды енгізу қажет екені мə. лімМұндай |
ақуызар мүшелерде жəне |
|||
ұлпаларда өте аз мөлшерде кездеседі, ал олардың |
кейбіреулерінде дəлме-дəл |
|||
ерекшелік |
қасиеттердің |
болатыны– |
қатаң ескеруді |
талап етеді. Оларды тек |
донорлық қандардан немесе өліктер материалдарынан алуға болады. Мұндай ақуыздарды микробтық синтез жолымен алу , тиімдіүшін технологиялық жағдайларда, өнеркəсіпте қолдануға арналған микроорганизмдерге бөгде гендерді
енгізу тəсілдерін жақсылап |
игеру жəне осындай микроорганизмдердің тиі |
|
қасиетін одан əрі жетілдіріп, үдерісті жеделдетуге тигізетін əсерін арттыра |
түсу |
|
керек. |
|
|
Соңғы кездерде нуклеин |
қышқылдарын алмастырудың түбегейлі |
əдісте |
жасалды. Соның негізінде молекулалық биология мен генетиканың жаңа бір саласы
– ген инженериясы қалыптасып дамуда. Ген |
инженериясы |
одан бұрынырақ |
||
қалыптасқан |
хромосоманы, |
генотипті |
өзгертуге |
қарағанда, ДНҚ-ны |
рекомбинациялау арқылы ген қызметіне араласуға мол мүмкіндіктер туғызды. Ген инженериясы жəне генетикалық инженерия дегендер егіз ұғымдар, дегенмен соңғысының жеке гендерден гөрі геномның ірі бөлшектеріне қатысы көбірек келеді.
3. Гендік инженерияны қолдану əдістері
Биолог зерттеушілері үшін бактериалды жасушаларының тұқым қу аппаратары үлкен қызығушылық тудырады. Бактерия жасушаларында ДНҚ-ы ядро орналасатын орта тұсында жайғасады. Бірақта, өсімдіктер мен жануарлар жасушаларындағы ядро құрамына кіретін ДНҚ-мен салыстырғанда, бактериялдық ДНҚ ядролық қабықпен қоршалмай, цитоплазмада бос күйінде жатады. Бактерия хромосомалары шеңберленіп біткен жіпшелерге ұқсас болады. Олар екі үзікті (цепь) ДНҚ молекулаларынан құралады.
Біздерге, ДНҚ |
молекуласының əрбір үзіктері бірнеше |
|
нуклеотидтерд |
|
тұратыны, яғни |
полинуклеотидті болып |
келетіні жəне |
өз |
кезегінде |
нуклеотидтер құрамына көміртегі дезоксирибозасы, фосфор қышқылдарының қалдықтары мен азот негізді заттар кіретіні белгілі. Нуклеотидтер бір-бірінен осы
азот негізді заттар түрлерінің кіруіне |
байланысты |
ерекшеленеді. Бұл заттар |
қатарында аденин, гуанин, цитозин, тимин |
болуы мүмкін |
екендігін еске түсіре |
13
кетейік. ДНҚ үзіктерін құрайтын нуклеотидтер бірбірімен фосфор қышқылы мен дезоксирибозалар арқылы белгілі бір ретпен байланысады. Мысалы, адениннің (А) бір үзігіне қарсы тиминннің(Т) екінші үзігі, ал гуанинге (Г) қарсы – цитозин (Ц) орналасады. Екі нуклеотидтер сутегілік байланыстар арқылы бірігіп тұрады жəне де мұнда олар бір-бірін толықтырып отырғандай болады. Сондықтан гуанинді – цитозинге, ал аденинді – тиминге комплементарлы (лат. Complemente – толықтыру деген мағынада) деп айтады.
ДНҚ молекуласындағы нуклеотидтердің белгілі бір ретпен орналасуы, олардағы қандай да бір ақуыздың синтезделуі жөніндегі информациясын сақтайды. Егерде, белгілі бір себептермен нуклеотидтердің біреуі кірмей немесе алмасып қалса, басқа
ақуыз синтезі жүре бастайды. Сондықтан |
ақуыз |
құрылымы жөнінде |
мəлімет |
сақталған ДНҚ бөлігі – геном деп аталады. |
|
|
|
Кейбір бактериялардың хромосомдар |
құрылымы толықтай белгілі болға. |
||
Мысалы сенна таяқшасы деген атпен белгілі бактерияларының хромосомасы200- |
|||
ден аса геннен тұрады. Ғалымдар осы |
гендердің хромосома |
шеңберін |
|
орналасуының картасын жасаған. |
|
|
|
Гибридті ДНҚ алу барысында атқарылатын ген |
инженериясы жұмыстарының |
||
дұрыс жүруіне, бактериалды жасуша бірліктеріне кіретін плазмидтер деп аталатын
құрылымдар |
үлкен |
роль |
атқаратындығы |
анықталғ. Плазмидан |
терминін |
||||
қолданысқа америкалық ғалым генетик жəне биохимик Джошуа Ледерберг1952 ж. |
|
||||||||
енгізді. |
|
|
|
|
|
|
|
|
|
Плазмидтер дегеніміз – құрамына 2250-ден 400000 жұпқа дейін нуклеотидтер |
|
||||||||
кіретін ДНҚ молекуласы болып табылады. Олар |
бактериялар |
жасушаларында |
|||||||
бірден |
бастап |
бірнеше |
ондық |
жұп |
көлемінде |
кездесуі |
мүм |
||
хромосомалардан бөлек орналасады. Бактерия хромосомасы сияқты плазмидтер де шеңбер тəрізді болып біткенімен, хромосомалардан əжептеуір кіші болады. Плазмидтердегі ДНҚ саны бактериялды хромосомалармен салыстырғанда20-1000
есеге дейін аз болады. Плазмидтер бактериялды |
хромосомаларға |
тəуелсіз екі |
еселене алу (репликациялану) ерекшеліктеріне |
ие. Плазмидтер |
бактериялды |
жасушаларының дəрілерге, мысалы антибиотиктерге қарсы тұру қабілеттіліктеріне жауап береді. Бұлардың ең маңызды қасиеттері қатарына бір жасушадан екінші жасушаға өте алу ерекшелігін жатқызуға болады.
Егерде плазмидаға ДНҚ-ның қандай да бір бөлігін ендірсе, ол плазмидамен бірге репликацияланады. Осы ерекшеліктің арқасында адамдарды қызықтыратын қажетті гендерді еселей көбейтіп (клондап) алуға болады.
Генетикалық инженерияның əдістерін дамытудаэндонуклеазалар деп аталатын спецификалық ферменттер өте үлкен маңызға ие болды. Ферменттер – ағзадағы жүретін биохимиялық үдерістердің қарқынын бірнеше есе арттыра алатын, шығу тегі ақуыздық негізді болып келетін биологиялық катализаторлар екенін еске түсіре кетейік. Эндонуклеазалар бактериялды жасушаларының өздерімен синтезделеді жəне шеңберлі ДНҚ молекулаларын сызықты бөліктерге кесе алатын қабілетке ие екендіктері анықталған. Осындай ферменттердің барлығы жөніндегі ойлар, өткен
ғасырдың |
50-ші |
жылдарының |
басында |
бірнеше |
зертханаларда |
жүргізі |
|
тəжірибелер |
барысында, бактериалды |
жасушаларында |
бактериофагтардың |
||||
(бактерия |
вирустары) көп көбейе |
алмауы себебі анықталғаннан |
кейін жасалынған |
||||
14
болатын. Рестрикция деген сөз бір нəрсенің шектелуін білдіреді. Сол себептен бактериофагтардың көбейуіне шектеу келтіретін фермент түрлері – рестриктазалар деп атала басталды. Қазіргі кезде оларды əртүрлі микроорганизмдерден бөліп алады. Олар генетикалық инженерияда жасалатын тəжірибелерде міндетті түрде
қолданыла |
бастады. Рестриктазалар |
көмегімен ДНҚ |
молекулаларын |
қажетт |
жерлерінен кесіп алу мүмкін болды. |
|
|
|
|
Бактериялды жасушаларда рестриктаза ферменттерімен бірге, рестриктазалар |
||||
кескен ДНҚ |
бөліктерін қайта«тіге» |
немесе «желімдей» |
алатын лигаза |
атты |
ферменттер де синтезделетіні белгілі болды. Сондықтан, рестриктаза мен лигаза ферменттерінің ашылуы генетикалық инженерияның дамуына үлкен жол ашты. Бұл ферменттердің көмегімен, қажетті қасиеттерге ие ДНҚ молекулаларын арнайы құрастыру мүмкіндігіне жол ашылды.
Ген инженериясының негізгі мазмұны – ағзадағы қажетті генді бөліп алып, оны вектормен біріктіруге саяды. Мұнда көбінесе генетикалық вектор ретін плазмидалық ДНҚ-ын пайдаланады. Осы үдеріс нəтижесінде алынған гибридті ДНҚ
молекуласы |
(рекомбинантты, |
яғни рДНҚ), |
бактерия жасушасына ендіріледі. |
Бактерия |
жасушасында |
векторлардың |
еселенуі(репликация) нəтижесінде, |
дайындалған гибридті рДНҚ молекулаларының да саны арта бастайды. Бұл үдеріс – рекомбинантты ДНҚ-ын клондау деп аталады.
Жалпылай алғанда гендік инженерия жұмыстарының барысы өз кезегіме атқарылатын келесідей кезеңдерден тұрады деуге болады:
1.Ағза жасушаларынан (Е. coli) қажетті ДНҚ-ын жəне ДНҚ векторларын бөліп алу.
2. |
ДНҚ-ын |
арнаулы |
рестриктаза |
|
ферменттері |
арқылы |
қажетті |
|
гені |
||||
|
бөліктерінен бөлу жəне бөлінген генді пайда |
болған |
рестриктаз |
||||||||||
|
қосындылар арасынан ажыратып алу. |
|
|
|
|
|
|
|
|||||
3. |
Ағзалардан |
алынған |
|
қажетті |
|
ДНҚ-ның |
бөліктерін(гендерді) лигаза |
|
|||||
ферменттерін қолдана отырып ДНҚ векторларымен біріктіру (рДНҚ құрау). |
|
|
|||||||||||
4. |
Құрастырылған рДНҚ-ын |
арнайы |
дайындалған |
ағзаларға, мысалы |
Е. |
coli |
|
||||||
немесе денелік жасушаларына ендіру. |
|
|
|
|
|
|
|
|
|||||
5. |
ДНҚ |
молекулалары егілген гибридті бактерияларды арнайы қоректік |
|||||||||||
ортаға |
себу |
арқылы, тек |
олардың |
көбеюін(репликациясын) |
қамтамасыз |
ету |
|||||||
(Гендердің генотиптік көрініс беруі экспрессия деп аталады). |
|
|
|
|
|
||||||||
6. |
ДНҚ |
молекулалары |
егілген |
гибридті |
бактериялар |
|
коло |
||||||
|
идентификациялау. |
|
|
|
|
|
|
|
|
|
|
||
7. |
Клондалған |
ДНҚ (клондалған гендерді) бөлу жəне оларды сипаттау. Бұл |
|
||||||||||
|
кезеңде |
клондалған |
|
ДНҚ |
бөліктеріндегі |
азотты |
негіздері |
де |
|||||
(секвенирование) тасталады.
Жоғарыда айтылған гендік инженерия жұмыстарының кезеңдерін жалпылай сызбанұсқа түрінде көрсететін болсақ, келесі келтірілген суреттегідей болады (сурет-3)
15
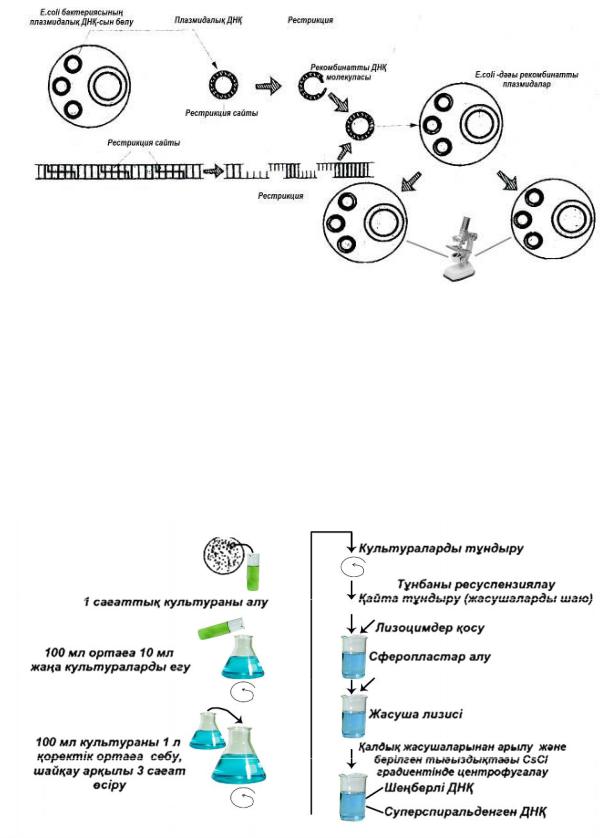
Клондалған ДНҚ бөлу жəне сипаттау
3-сурет. Гендік инженерия жұмыстарының атқарылу кезеңдері
Қажетті ДНҚ-ын бөліп алу жұмыстары техникалық жағынан қандай да бір қиыншылықтар тудырмайды. Бұл əдіс ғылымда40 жылдай қолданылып келеді.
Осындай экстракциялау жұмыстары нəтижесінде жасушалар мен |
ұлпалар |
құрамында ақуыз немесе басқа да заттары жоқ ДНҚ-ның таза түрі алынады. Қажетті |
|
ДНҚ-ын бөліп алу жұмыстарының атқарылу кезеңдерін төменде |
келтіріл |
сызбанұсқа түрінде көрсетуге болады (сурет-4). |
|
4-сурет. Қажетті ДНҚ-ын бөліп алу жұмыстарының атқарылуы
ДНҚ-ын рестриктаза ферменттері арқылы бөліктерге бөлужұмыстарын бірнеше əдістер арқылы жүзеге асыруға болады. Бірақта олардың барлығын гендік
16
инженерия мақсатында қолдана алмаймыз. Ол үшін үлескілерге бөлінген ДНҚ-ның қайтадан бір-бірімен біріге алатындай күйде болуын қамтамасыз ету қажет болады.
Өткен ғасырдың 70 жылдарының орта шенінде рестриктаза ферменттерінің көмегімен ДНҚ-ын кесілген ұштарына ДНҚ-ның екінші бір бөліктерін жапсыра алатындай етіп бөлу мүмкіндігі ашылғаннан кейін, генетикалық инженерияның дамуына кең жол ашылып, рекомбинантты ДНҚ алу технологиясыны қалыптасуына бастау болды. Мысалы E. coli ферменті алты жұп нуклеотидтен тұратын ДНҚ үлескілерін белгілеп, осы бөліктегі ДНҚ-ның екі жіпшелі құрылымын олардың кесілген екі ұштарының əрқайсысында төрт нуклеотидтерден құралған бір жіпшелі үзіктерден тұратындай етіп кесе алады .екенКесілген мұндай ұштар
комплиментарлы жəне бір-бірімен сутегілік байланыс арқылы |
өзара əрекеттес |
||
алатын болғандықтан «жабысқақ» деп |
аталады. Осы |
мақсатта |
қолданылатын |
ферменттер, ДНҚ үлескілерін олардың |
құрамындағы |
нуклеин |
қышқылдарының |
шығу тегіне байланыссыз танып-белгілей беретін болғандықтан, ДНҚ бөлінген үлескілері ұштарының барлығы бірдей«жабысқақ» күйде болады. Сонымен бірге, өздерінің əрекеттері нəтижесінде ДНҚ кесілген үлескілеріндегі екі үзік ұштары да жапсырылған, яғни «доғал» күйінде болып шығуы себепті, бір-бірімен жабыса алмайтындай дəрежеде дайындайтын рестриктаза ферменттерінің де бар екендігі белгілі болды.
Рестриктаза ферменттерінің ДНҚ-ын кесу нəтижесінде олардың көптег бөліктері алынады. Бөліктердің саны бұл мақсатта қолданылатын рестриктаза ферменттерінің түріне байланысты болады. Гендер «кітапханасын» жасау үшін, ДНҚ-ын бірнеше ондаған немесе жүздеген мың бөліктерге бөлу қажет болады.
4.1 Рекомбинантты ДНҚ молекуласын конструкциялау
Бактериалды жасушаға ендірер алдында, молекулалық деңгейде клондауға арналған ДНҚ сегментінің (геннің) репликацияға қабілетті, яғни репликонды болуы
қажет. Алайда |
ол мұндай |
қабілетке |
ие . емесСондықтан олардың |
жасушаға |
|
тасымалдануы мен жасушадағы клондалған гендердің белгілі |
болып тұр |
||||
қамтамасыз ету үшін, оларды генетикалық векторлармен біріктіреді. |
|
||||
Векторлар |
дегеніміз – |
гендерді |
пробиркадан |
биологиялық |
зерзатын |
тасымалдау жəне оларда қалыпты қызмет етулерін қамтамасыздандыруға қажетті генетикалық құрылым. Вектор ретінде плазмидалар мен фагтар бола алатындықтары жөнінде біздер білеміз. Плазмидалардың қандай да бір антибиотиктерге резистентті болуы бактерияларды осындай антибиотиктерге селекция жүргізуге мүмкінд тудырады жəне рекомбинантты ДНҚ молекуласының оңай анықталуы себепті, олар жақсы векторлар болып саналады.
Рекомбинантты ДНҚ молекуласын конструкциялауды, зерттелетін ДНҚ мен векторлық ДНҚ рестрикталарын алғаннан кейін бастау қажет болады. Мұның мəні – зерттелетін ДНҚ рестрикт сегментін(бөлшегін) векторлық ДНҚ рестриктасымен қосуға саяды жəне соның нəтижесінде олардың пішіні шеңберлі құрылымна, ұштары «жабысқақ» болып келетін түзу сызықты ДНҚ бөліктеріне айналады.
Кейіннен плазмидалық ДНҚ бөліктері лигаза ферменттерінің əсерімен сырттан əкелінген ДНҚ бөліктерімен қосылады. Нəтижесінде плазмида қайтадан шеңберлі
17
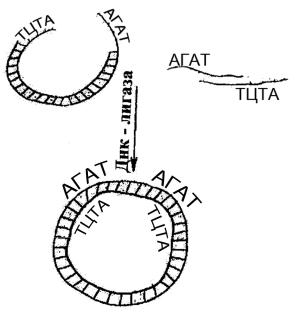
құрылымына ие болады. Мұндай жұмыстарда көптеген плазмидалар пайдалануы себепті, құрамында əртүрлі гендері бар ДНҚ комбинациялары түзілуі мүмкін (сурет- 5).
5-сурет. ДНҚ-ын біріктіру (лигирование)
Көптеген рестриктазалар, мысалы Eco R-І сияқтылар төрт негізден тұратын «жабысқақ» ұштыларды шығарады. Осындай төрт негізден тұратын«жабысқақ» ұштыларды лигирлеу төменгі температураларда (120С дейін) жүреді.
Егерде рестрикциялау барысында «жабысқақ» ұшты бөлшектер пайда болмаса, оларды трансфераза ферменттерін қолдана отырып«жабысқақ» ұштыларға «мəжбүрлі» түрде айналдырады. Бұл фермент ДНҚ-ның 31 ұшына нуклеотидтерді қосады. Бір бөлшегіне поли-А құйрықшасы жалғанса, екіншісіне – поли-Т құйрықшасы жалғанады. Қажетті ұшты ДНҚ-ын алу үшін– полимеразды тізбектік реакциясы (ПТР немесе орысшасы – полимеразная цепная реакция – ПЦР) деген де қолданылады. ПТР-ның мəні – жасушадан бөлініп алынған ДНҚ-ын денатурациялау мен «күйдіру» жəне ДНҚ олигонуклеотидтерінің əрқайсысы 15-20 нуклеотидтерден тұратын ренатурацияланатын үзіктеріне жалғау болып табылады.
3.2 Рекомбинантты ДНҚ молекуласын (рДНҚ) жасушаға ендіру
Біріктірілген рДНҚ-ын арнайы дайындалған жасушаларға ендіру– гендік инженериялық манипуляциялау жұмыстарының соңғы кезеңі болып табылады. Мұндағы ең күтілетін нəтиже – ендірілген жасуша құрамында олардың көбейуі мен өздерінің қызметтерін дұрыс атқаруы, яғни қажетті ақуыз синтезін қамтамасыз етуі керек. Осындай жасуша ретінде, хромосомалар құрамы жақсы зерттелген ішек таяқшасы (Е. coli) бактериялары жиі қолданылады. Бактериялды жасушалары рекомбинантты ДНҚ-ын қабылдауға лайық болуы үшін алдын-ала кальций немесе рубидиймен өңделеді.
Жасушаларға ДНҚ-ның кіру мүмкіндігін арттыру мақсатында электропорация əдісі жиі қолданылады. Бұл əдіс бойынша жасушаларға интенсивті электр өрісімен əсер ету жүзеге асырылады. Осындай əсер ету нəтижесінде жас
18
мембраналарында қуыстар пайда ,болыпжасушалардың ДНҚ-ын жақсы қабылдауына мүмкіндіктер туылады. Құрамында ДНҚ бөліктері бар плазмидалар ішек таяқшасы жасушасына енеді. Құрамына көп ДНҚ бөліктері бар плазмида енген соң, ішек таяқшасы бактериялары əртүрлі гендермен байытылады.
Кейіннен, рекомбинантты ДНҚ молекуласы бар бактериялары, осындай қажетті
жасушалардың (рДНҚ-лы) селекциясын |
қамтамасыз |
ету үшін |
керек |
болатын, |
|||
арнайы |
антибиотиктермен |
байытылған, ЕПА |
(етті-пептонды |
агар) қоректік |
|||
ортасына |
себіледі. Трансформация |
жиілігі |
көп |
болмайды. Көбінесе |
бір |
||
трансформант 105 себілген жасушаларынан пайда болады. Егерде вектордың тегі фагтан шықса, жасуша трансфекциясын (бактериялар немесе ашытқылар) фагпен жүргізеді.
Жануарлардың денелік (сомалық) жасушаларына келетін болсақ, олардағы ДНҚ трансформациясын, плазмалық мембранадан ДНҚ-ның тез өтуін қамтамасыз ететін арнайы химиялық заттардың көмегі арқылы жүзеге асырады. Сонымен бірге құрбақа ооциттеріне, қоректік ортада өсіріліп жатқан денелік(сомалық) жасушаларына немесе сүт қоректілердің эмбриондарына рДНҚ-ын тікелей (иньекциялауегу) жолымен ендіруге де болады.
Қоректік (селективтік) ортада арнайы өсірілген трансформант колониялары, қажетті геномның немесе кДНҚ бөлшектері(гені) бар жасушалар жиынтығы болып табылады. Осындай клондар коллекцияларын ДНҚ кітапханасы деп те атайды жəне олар гендік-инженерлік жұмыстарында кеңінен қолданылады.
Гендерді клондау жұмыстарының соңғы кезеңдерінде, қажетті гендері бар бактериялар түрлі əдістер арқылы бөлініп алынады жəне тексеріліп болғаннан соң, құрамында рДНҚ-сы бар, қажетті ақуызды синтездеуді бақылай алатын құнды колониялары ары қарай пайдаланылады.
Вектор ретінде плазмидалармен қатар əртүрлі бактериофагтар, хайуандар мен өсімдіктер вирустарын да пайдалануға болады. Алайда, хайуандар мен өсімдіктер жасушаларында хромосомалар құрамы мен гендер əрекеттері бактериялардан бірталай айырмашылықтары болуы себепті, бұларда гендікинженериялық манипуляциялар жүргізу жұмыстары қиынырақ келеді.
3.3 Гендік инженерияның практикалық маңызы |
|
||||
Молекулалық |
генетика |
мен |
генетикалық |
инженерияның |
қол |
табыстарының арқасында, жасушаға сырттан жаңа генетикалық информация енгізу арқылы организмдердің алдын-ала жобаланған тұқым қуалайтын бағдарламасын
жасауға |
болады. Ондай |
информацияның қолдан |
синтезделінуі мүмкін |
немесе |
|
табиғи генетикалық зат |
ретінде əртүрлі организмдерден бөлініп алынады. Сөйтіп, |
||||
тəжірибелік (эксперименттік) əдіспен, табиғи-эвюлюциялық |
жолмен пайда |
бола |
|||
алмайтын жаңа генетикалық жүйе жасалады. |
|
|
|
||
Гендік |
инженерия |
жетістіктері– денсаулық |
сақтау, |
ауылшаруашылығы, |
|
биотехнология жəне микробиологиялық өндіріс салаларында аса зор практикалық
мəні бар |
проблемаларды шешуде маңызды роль атқарады. Гендік инженерия |
|
əдістерін |
жетілдіру |
денсаулық сақтау мен ауылшаруашылығы үшін қаж |
гормондар, |
ферменттер |
жəне антибиотиктерді синтездейтін микроорганизмдердің |
19