

Экспрессия генов
В этом разделе рассмотрена экспрессия генов и её регуляция. Экспрессия генов (рис. 3–13) происходит в несколько этапов: транскрипция процессингтрансляцияпосттрансляционная модификация. На каждом из этих этапов на макромолекулы в виде гена, первичного транскрипта, мРНК, полипептида и белка воздействуют различные регуляторные факторы.

Рис.3–13.Этапыэкспрессиигена[2]. Направление процессов указано стрелками, этапы экспрессии гена названы слева; участники этапов экспрессии (ДНК, транскрипт, мРНК, полипептид, белок) пронумерованы римскими цифрами справа. Транскрибируемый участок гена ограничен цифрами 1 (сайт инициации транскрипции) и 2 (сайт терминации транскрипции), реализуемые в полипептиде участки гена и мРНК («рамка считывания») — цифрами 3 и 4. В составе мРНК указан полиА-«хвост».
Транскрипция
Транскрипция — первый этап реализации генетической информации — приводит к появлению на матрице ДНК первичного транскрипта (РНК), состоящего из линейной последовательности транскрибированных интронов и экзонов. ДНК служит только матрицей для синтеза РНК и в ходе транскрипции не изменяется. В основе механизма транскрипции (полимеразная реакция, катализируемая РНК–полимеразами) находится принцип комплементарного спаривания оснований. Синтез РНК начинается в определённых последовательностях (сайтах) ДНК — промоторах и завершается в сайтах терминации.
Транскриптон— участок ДНК между промотором и сайтом терминации — единица транскрипции. Каждый транскриптон содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные факторы транскрипции.
Факторытранскрипции— белки, взаимодействующие с определёнными регуляторными сайтами ДНК, инициирующие, ускоряющие, замедляющие или блокирующие транскрипцию конкретного гена.
РНК-полимеразы. Биосинтез РНК осуществляют ДНК–зависимые РНК–полимеразы: РНК–полимераза I синтезирует пре-рРНК, РНК–полимераза II — пре-мРНК, РНК-полимераза III — пре-тРНК. РНК–полимеразы состоят из нескольких СЕ, СЕ(сигма) выполняет регуляторную функцию, один из факторов инициации транскрипции.
Стадиитранскрипции. В ходе транскрипции выделяют 3 стадии: инициацию, элонгацию и терминацию.
Инициация. Промотор активирует фактор инициации — большой белок — ТАТА-фактор (взаимодействует со специфической последовательностью нуклеотидов промотора — ТАТААА [ТАТА-бокс]). Факторы инициации вызывают изменение конформации РНК–полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, в которой матрица доступна для синтеза цепи РНК. После синтеза олигонуклеотида из 8–10 нуклеотидных остатков,‑субъединица РНК–полимеразы отделяется от фермента, а её место занимают несколько факторов элонгации.
Элонгация. Факторы элонгации облегчают расхождение цепей ДНК. По мере продвижения РНК–полимеразы по матрице ДНК в направлении от 3`- к 5`-концу впереди неё происходит расхождение, а позади — восстановление двойной спирали ДНК.
Терминация. Раскручивание двойной спирали ДНК в области сайта терминации транскрипции делает его доступным для фактора терминации. Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице ДНК, и РНК-полимеразы от матрицы ДНК. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы.
Процессинг
Первичные транскрипты подвергаются ряду ковалентных модификаций (модификации концов пре-мРНК и сплайсинг), в результате осуществления которых формируется способная к трансляции зрелая мРНК.
Модификация5`-конца, иликэпирование. Катализирует кэпирование гуанилилтрансфераза, гидролизующая макроэргическую связь в молекуле ГТФ и присоединяющая нуклеотиддифосфатный остаток к 5`-концу синтезированного фрагмента РНК с образованием 5`,5`-фосфодиэфирной связи. Модифицированный 5`-конец обеспечивает инициацию трансляции, удлиняет время жизни мРНК и защищает её от действия 5`-экзонуклеаз в цитоплазме.
Модификация3`-конца. ПолиА-полимераза формирует полиА-последовательность (полиА-«хвост»), состоящий из 100–200 остатков адениловой кислоты.
Сплайсинг. Последовательности нуклеотидов, присутствующие в ДНК, но не входящие в состав мРНК, названы некодирующими (интроны), а последовательности, присутствующие в мРНК, — кодирующими (экзоны). Последовательности интронов «вырезаются» из первичного транскрипта, а концы экзонов соединяются друг с другом. Такую модификацию РНК называют «сплайсинг» (от англ.tosplice— сращивать). Процесс «вырезания» интронов протекает при участии малых ядерных рибонуклеопротеинов. Сплайсинг происходит в ядре, а в цитоплазму поступает «зрелая» мРНК. Гены человека содержат больше интронов, чем экзонов, поэтому очень длинные молекулы пре-мРНК (около 5000 нуклеотидов) после сплайсинга превращаются в более короткие молекулы цитоплазматической мРНК (от 500 до 3000 нуклеотидов).
Трансляция
Для синтеза полипептидной цепи необходимы следующие основные компоненты: аминокислоты, тРНК (около 50 разных тРНК), мРНК, рибосомы, источники энергии (АТФ и ГТФ), белковые факторы инициации, элонгации и терминации трансляции. Процесс трансляции схематически отображён на рис. 3–14.
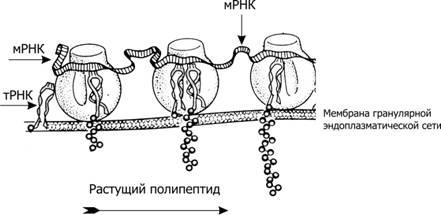
Рис.3‑14.ТрансляциямРНК[9]. Рибосома (80S) состоит из большой (внизу) и малой (вверху) СЕ. Большая СЕ (60S) содержит 5S, 28S и 5,8S РНК и до 50 белков, малая СЕ (40S) включает 18S РНК и более 30 белков (S — константа седиментации). Рибосома имеет три различных участка связывания РНК — один для мРНК и два для тРНК. Пептидил-тРНК-связывающий участок удерживает молекулу тРНК, присоединённую к растущему концу полипептидной цепи; расположенный рядом аминоацил-тРНК-связывающий участок (акцепторный) фиксирует только что поступившую в рибосому молекулу тРНК с аминокислотой. тРНК называют «адапторные молекулы», так как к акцепторному их концу присоединяется определённая аминокислота, а с помощью антикодона они узнают специфический кодон на мРНК. Другими словами, тРНК адаптируют разные по структуре и расположению в составе макромолекул РНК и белка мономеры РНК (нуклеотиды) и белка (аминокислоты). СЕ рибосомы выполняют разные функции: малая СЕ присоединяет мРНК и декодирует информацию с помощью тРНК, большая СЕ ответственна за образование пептидных связей.
Генетическийкод. Трансляция информации, записанной в мРНК (последовательность нуклеотидов), в аминокислотную последовательность полипептида требует наличия генетического кода, т.е. правил, согласно которым первичная структура нуклеиновой кислоты (чередование четырёх нуклеотидов) в ДНК и мРНК определяет первичную структуру белка (специфическая последовательность аминокислот в полипептидной цепочке). Другими словами, необходим своего рода «алфавит».
Кодоны. Буквами «алфавита» являются кодоны —триплетынуклеотидов, кодирующие конкретную аминокислоту. Получается, что суммарное количество кодирующих последовательностей из четырёх нуклеотидов по три (кодон) равно 64 (43=64), т.е. более чем в три раза превышает минимальное количество кодонов, необходимое для кодирования 20 аминокислот.
Свойствагенетическогокода. Для генетического кода характерны специфичность и вырождённость (избыточность).
Специфичность. Каждому кодону соответствует только одна конкретная аминокислота. Так, кодон GGU кодирует аргинин, кодон AUA — метионин, кодон CCA — пролин (U — урацил, C — цитозин, A — аденин, G — гуанин).
Вырождённость— включение в полипептидную цепочку одной и той же аминокислоты кодируют несколько кодонов. Так, аргинин кодируют 6 триплетов (CGU, CGC, CGA, CGG, AGA и AGG), пролин — 4 (CCU, CCC, CCA, CCG), но метионин — только 1 триплет AUA.
Синтезполипептиднойцепипроисходит от N- к С-концу при чтении генетического кода от 5`- к 3`-концу мРНК. После освобождения 5`-конца мРНК к этому концу может присоединиться новая рибосома, на которой начинается рост ещё одной полипептидной цепи. Как правило, с одной и той же мРНК одновременно связано много рибосом (полирибосома, или полисома).
Этапытрансляции— инициация, элонгация и терминация.
Инициация. На этом этапе трансляции участвуют факторы инициации eIF (от англ.eukaryoticinitiationfactors). После образования комплекса, состоящего из тРНК, мРНК, рибосомы и факторов инициации трансляции (см. рис. 3–14), СЕ 40S рибосомы скользит по некодирующей части мРНК до тех пор, пока не достигнет инициирующего кодона AUG («3» на рисунке), после чего СЕ останавливается и связывается с другими факторами инициации.
Элонгация. Рибосома с помощью аминоацил–тРНК последовательно «читает» кодоны мРНК, следующие за инициирующим кодоном в направлении от 5` к 3`-концу, наращивая полипептидную цепочку за счёт последовательного присоединения аминокислот.
Терминациятрансляции наступает по достижении одного из стоп-кодонов (UAG, UAA или UGA, цифра «4» на рисунке), для которых нет соответствующих тРНК. Вместо тРНК к рибосоме присоединяются 2 факторы терминации. Один из них катализирует гидролитическое отщепление синтезированного пептида от тРНК, а другой вызывает диссоциацию рибосомы на СЕ.