

Клеточный цикл
Клеточный цикл (рис. 3–11) cостоит из митоза (М‑фаза, рис. 3–12) и интерфазы. В интерфазе последовательно различают фазы G1, S и G2.
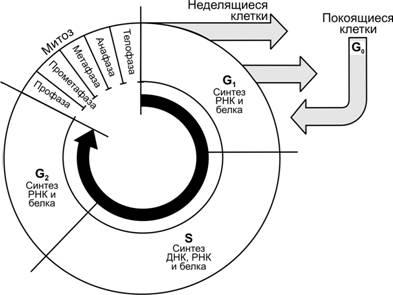
Рис.3–11.Стадииклеточногоцикла[11]. В клеточном цикле различают митоз — сравнительно короткую фазу M и более длительный период — интерфазу. Фаза M состоит из профазы, прометафазы, метафазы, анафазы и телофазы; интерфаза складывается из фаз G1, S и G2. Клетки, выходящие из цикла, более не делятся и обычно специализируются. Клетки в фазе G0обычно не возвращаются в цикл.
Интерфаза
G1— обычно самая продолжительная фаза цикла, следует за телофазой митоза. В эту фазу клетка синтезирует РНК и белки. Продолжительность фазы — от нескольких часов до нескольких дней. У быстро делящихся клеток (эмбриональные и неопластические) эта фаза непродолжительна.
G0. Клетки могут выйти из цикла и находиться в фазе G0. В фазе G0клетки начинают дифференцироваться, достигая состояния терминальной (окончательной) дифференцировки (например, нейроны).
S. В фазу S в клетке продолжается синтез белка, происходит репликация ДНК. В большинстве клеток фаза S длится 8–12 ч.
G2. В фазу G2 продолжается синтез РНК и белка (например, синтез тубулина для микротрубочек митотического веретена), накапливается АТФ для энергетического обеспечения последующего митоза. Эта фаза длится 2–4 ч.

Рис.3–12.Митоз— M-фаза клеточного цикла [11]. После фазы G2клеточного цикла начинается M-фаза (митоз). Эта фаза состоит из пяти стадий деления ядра (кариокинез) и деления цитоплазмы (цитокинез). M-фаза заканчивается к началу фазы G1следующего цикла.
Митоз. В ходе митоза делятся ядро (кариокинез) и цитоплазма (цитокинез). Фазы митоза: профаза, прометафаза, метафаза, анафаза, телофаза (рис. 3–12).
Профаза. Каждая хромосома состоит из двух сестринских хроматид, соединённых центромерой, исчезает ядрышко. Центриоли организуют митотическое веретено. Пара центриолей входит в состав митотического центра, от которого радиально отходят микротрубочки. Сначала митотические центры располагаются вблизи ядерной мембраны, а затем расходятся, и образуется биполярное митотическое веретено. В этом процессе участвуют полюсные микротрубочки, взаимодействующие между собой по мере удлинения.
Прометафаза. Ядерная оболочка распадается на мелкие фрагменты. В области центромер появляются кинетохоры, функционирующие как центры организации кинетохорных микротрубочек. Отхождение кинетохор от каждой хромосомы в обе стороны и их взаимодействие с полюсными микротрубочками митотического веретена — причина перемещения хромосом.
Метафаза. Хромосомы располагаются в области экватора веретена. Образуется метафазная пластинка, в которой каждая хромосома удерживается парой кинетохоров и связанными с ними кинетохорными микротрубочками, направленными к противоположным полюсам митотического веретена.
Анафаза— расхождение дочерних хромосом к полюсам митотического веретена со скоростью 1 мкм/мин.
Телофаза. Хроматиды подходят к полюсам, кинетохорные микротрубочки исчезают, а полюсные продолжают удлиняться. Образуется ядерная оболочка, появляется ядрышко.
Цитокинез— разделение цитоплазмы на две обособляющиеся части. Плазмолемма втягивается между двумя дочерними ядрами в плоскости, перпендикулярной длинной оси веретена. Борозда деления углубляется, и между дочерними клетками остаётся мостик — остаточное тельце. Дальнейшее разрушение этой структуры приводит к полному разделению дочерних клеток.
Пролиферация клеток, происходящая путём митоза, жёстко регулируется множеством молекулярных сигналов. Регуляторы клеточного цикла и митоза подразделяют на внутриклеточные и межклеточные.
Внутриклеточныемолекулярныесигналымногочисленны, среди них в первую очередь следует назвать собственно регуляторы клеточного цикла (циклины, циклин–зависимые протеинкиназы, их активаторы и ингибиторы) и онкосупрессоры.
Циклины,циклин–зависимыепротеинкиназы(Cdk),активаторыиингибиторыциклиновиCdk. Скоординированная деятельность этих многочисленных регуляторов клеточного цикла обеспечивает как переход клеток от фазы к фазе клеточного цикла, так и точное выполнение событий каждой фазы. Нарушения скоординированной деятельности приводят как к искажениям прохождения клетки по фазам клеточного цикла (патология митоза), так и к потере контроля за пролиферативными потенциями клетки, что может привести к бесконтрольной её пролиферации. Другими словами, происходитформированиенеконтролируемогоклона, т.е.опухолевыйрост. Клетки таких клонов называют трансформированными, или малигнизированными. Главная причина появления пролиферативно неконтролируемых клеток — мутации генов, кодирующих структуру собственных регуляторов клеточного цикла.
Онкосупрессорыв нормальной клетке постоянно контролируют множество процессов, в том числе и пролиферацию клеток (точнее, онкосупрессоры блокируют клеточный цикл). Известно множество онкосупрессоров (например, Rb, p27, p53, PTEN), среди которых p53 известен как главный супрессор развития опухоли. Мутации генов онкосупрессоров неизбежно приводят к появлению бесконтрольно пролиферирующего клеточного клона.
Межклеточныесигналы. Регуляция клеточного цикла и пролиферация (или блок пролиферации) клеток регулируются внеклеточными сигналами в виде гормонов (например, гормон роста, эстрогены, ФСГ), цитокинов (например, ИЛ и интерфероны), факторов роста (например, фактор роста эпидермиса). Некоторые из таких молекулярных сигналов расцениваются как стимулирующие митогенную активность факторы (митогены). Существенное влияние на пролиферативную активность клеток имеют и контакты с элементами межклеточного матрикса (например, с ламинином и фибронектином). После взаимодействия с соответствующими рецепторами такие митогенные сигналы (в значительном числе случаев при помощи связанных с G‑белками протеинкиназ) передаются на соответствующие участки генома, активируя транскрипцию контролирующих фазы клеточного цикла генов (например, циклин–зависимых протеинкиназ). Взаимодействие лигандов с рецепторами и дальнейший внутриклеточный перенос информационного сигнала рассмотрены в главе 4 («Межклеточные взаимодействия»).