

Промежуточные нити
Промежуточные нити (филаменты) состоят из белков, специфичных для определённых клеточных типов; имеют диаметр 8–11 нм (табл. 3–1). Различают цитокератины эпителия, десмины мышц, виментины мезенхимы, ламины ядерной оболочки, а также нейрофиламенты, глиофиламенты и чётковидные волокна хрусталика. Промежуточные нити создают внутриклеточный каркас, обеспечивают упругость клетки, поддерживают упорядоченность расположения компонентов цитоплазмы. Иммуноцитохимические реакции с АТ конкретных типов промежуточных нитей нашли применение в цитодиагностике генеза опухолей.
Таблица 3‑1.Белки промежуточных филаментов (нитей) различных клеток
|
Филаментыибелки |
Функции |
|
Цитокератины кератиновых нитей эпителиальных клеток |
Создают тянущее усилие, взаимодействуют с десмосомами. Маркёры опухолей эпителиального происхождения. |
|
Десмины мышечных тканей |
Образуют внутриклеточный каркас, взаимодействуя с сократительными структурами |
|
Виментины клеток мезенхимного генеза |
Маркёры опухолей соединительной ткани |
|
Глиальный фибриллярный кислый белок глиофиламентов в астроцитах |
Маркёры астроцитом |
|
Белки нейрофиламентов нервных клеток |
Поддерживают форму отростков нейрона |
|
Факинин и филензин чётковидных волокон хрусталика |
Поддерживают структуру хрусталиковых волокон |
|
Ламины ядерной пластинки ядерной оболочки |
Организуют ядерную пластинку и лежащий около неё хроматин. |
Депо кальция
Цистерны гладкой эндоплазматической сети многих клеток специализированы для накопления в них Ca2+путём постоянного откачивания Ca2+из цитозоля, где нормально содержание Ca2+находится на уровне 10–7М. Подобные депо кальция функционируют в любой клетке (гладкая эндоплазматическая сеть). Особенно велика роль кальциевых депо в скелетной и сердечной мышцах, нейронах, хромаффинных клетках, яйцеклетке, эндокринных клетках и т.д. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на функции клеток и их пролиферацию путём изменения концентрации в цитозоле внутриклеточного посредника — Ca2+. Например, условием сокращения мышечных элементов является резкое повышение концентрации Ca2+в цитозоле. Для этого необходимо постоянно откачивать ионы кальция из цитозоля и накапливать их в специальных депо, образованных Ca2+-запасающими цистернами гладкой эндоплазматической сети (их название в мышечных элементах — саркоплазматический ретикулум). Внутри цистерн находятсяCa2+-связывающиебелки(кальсеквестрин, кальретикулин и др.) В мембрану цистерн–депо Ca2+встроеныCa2+‑насосы(Ca2+‑АТФаза), постоянно закачивающие Ca2+внутрь цистерн, иCa2+‑каналы, через которые происходит выброс Ca2+из депо при поступлении сигнала (рис. 3–10).
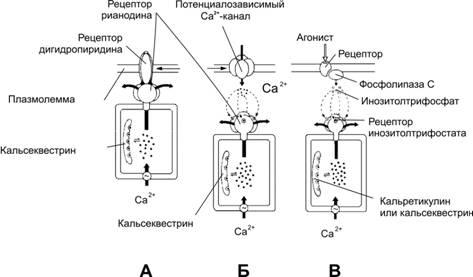
Рис.3–10.МеханизмвыбросаCa2+изцистернгладкойэндоплазматическойсетивсегда осуществляется при взаимодействии макромолекулярных структур плазмолеммы и стенки кальциевых депо.А—рецептор рианодинав мембране цистерны связан с Ca2+‑каналом, активируется дигидропиридиновым рецептором плазмолеммы;Б— рецептор рианодина в мембране цистерны непосредственно не связан с белками плазмолеммы, но активируется Ca2+при незначительном повышении концентрации этого катиона в цитозоле за счёт поступления в клетку через потенциалозависимые Ca2+‑каналы плазмолеммы;В— Ca2+освобождается из цистерн при активации рецепторов инозитолтрифосфата. Этот второй посредник образуется при участии фосфолипазы C в ответ на взаимодействие агониста со своим рецептором в плазмолемме. [11].
Известно несколько типов Ca2+‑каналов, в т.ч. управляемые рецепторами рианодина и вторым посредником (инозитолтрифосфат).
Рецепторырианодинаактивируются двояко: через рецепторы дигидропиридина и через потенциалозависимые Ca2+‑каналы плазмолеммы.
Рецепторыдигидропиридинавстроены в плазмолемму и реагируют на изменения её МП: изменение конформации рецепторов дигидропиридина активируетрецепторы рианодинаи приводит к выбросу Ca2+из депо. Подобный механизм функционирует в саркоплазматическом ретикулуме волокон скелетной мышцы.
Изменениямембранногопотенциалаоткрывают потенциалозависимые Ca2+‑каналы плазмолеммы, в результате в клетке несколько повышается концентрация Ca2+. Этот Ca2+активируетрецепторы рианодина, и запасённый в депо Ca2+выбрасывается в цитозоль. Этот механизм функционирует в нейронах и кардиомиоцитах.
Инозитолтрифосфат. Каналы в мембране цистерн открываются под действием инозитолтрифосфата (рис. 2‑10), образующегося при воздействии на клетку внешнего сигнала. Например, при взаимодействииангиотензиновсо своими рецепторомами в плазматической мембране ГМК активируется фосфолипаза С, и образуется инозитолтрифосфат. Последний диффундирует в цитоплазме, связывается с рецепторами инозитолтрифосфата в мембране цистерн гладкой эндоплазматической сети и открывает Ca2+‑каналы. Этот механизм функционирует в овоцитах, лимфоцитах, ГМК и других клетках.
Этанол—ингибиторрецепторовинозитолтрифосфата— участвует в развитии атаксии при алкогольной интоксикации. КлеткиПуркиньемозжечка имеютрецепторы рианодинаи инозитолтрифосфата в мембране цистерн, локализованных как в теле клетки (перикарион), так и в дендритах. Но в цистернах концевых разветвлений дендритов (шипики) присутствуют только рецепторы инозитолтрифосфата.