

Стероидогенез
Синтез гормонов коры надпочечника (из железы выделено не менее 50 стероидов) по-разному происходит в отдельных зонах коры. Стероидные гормоны и промежуточные метаболиты синтезируются на базе холестерола (C27H46O, рис.), поступающего в эндокринные клетки путём опосредованного рецепторами эндоцитоза из циркулирующих в крови ЛПНП. Незначительное количество холестерола синтезируется в самих эндокринных клетках. Холестерол аккумулируется в виде его эфиров в липидных каплях. Холестерол и все стероидные гормоны построены на основе 17-углеродной структуры — циклопентанпергидрофенантрена.

Рис. Циклопентанпергидрофенантрен (слева) и холестерол (справа) [11]. Слева указана нумерация колец и углеродных атомов в составе колец, справа — нумерация атомов углерода в составе боковой цепи. Тестостерон не образуется в коре надпочечников.
Стероидогенез обеспечивает множество ферментов, расположенных в митохондриях, цитозоле и гладкой эндоплазматической сети. Подавляющее большинство превращений в коре надпочечников, осуществляют ферменты, относящиеся к группе цитохромов P450. Ферменты этой группы, а также не относящейся к цитохромам 3b-гидроксистероид дегидрогеназы активируют АКТГ и ангиотензин II.
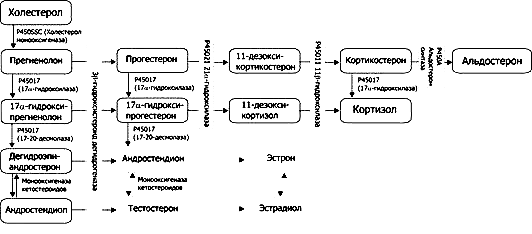
Рис. Пути синтеза стероидных гормонов надпочечника. Выделенные двойным контуром превращения в норме происходят вне надпочечника.
Мутации генов, кодирующих ферменты стероидогенеза, приводят к развитию врождённой дисфункции коры надпочечников — группе патологических процессов, сочетающихся с гиперплазией (увеличением массы) коры надпочечников.
Недостаточность 21-гидроксилазы — наиболее частая причина гиперплазии коры надпочечников (>90%). При этом дефицит кортизола стимулирует выработку АКТГ, что приводит к гиперплазии коры надпочечников и избыточной продукции андрогенов. Подобные нарушения при развитии плода часто вызывают изменения гениталий у девочек. При избытке андрогенов в постнатальном периоде происходит вирилизация (появление вторичных половых признаков мужского пола) в препубертатном возрасте и у молодых женщин. У младенцев мужского пола следствие избытка андрогенов во время развития плода — макрогенитосомия. В постнатальном периоде наступает преждевременное половое созревание. При тяжёлой (натрий-дефицитной) форме недостаточности — наряду с уменьшением синтеза кортизола — снижена продукция альдостерона; дефицит минералокортикоидов приводит к гипонатриемии, гиперкалиемии, дегидратации и гипотензии.
Недостаточность 17-гидроксилазы приводит к гиперсекреции стероидных метаболитов - развивается артериальная гипертензия. Поскольку фермент необходим и для синтеза половых стероидных гормонов наблюдается дефицит андрогенов и эстрогенов. Такие нарушения вызывают развитие гермафродитоподобных гениталий у мальчиков и первичную аменорею у женщин.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Состоит из двух долей, ткань состоит из фолликулов. В клетках щитовидной железы происходит синтез двух химически и функционально разных классов гормонов — йодированных – тироксин, трийодтиронин (синтезируются в эпителиальных фолликулах железы) и неиодированных – продуктов экспрессии кальцитониновых генов – тиреокальцитонин (синтезируются в так называемых светлых клетках — C‑клетки — фолликулов).
Йодсодержащие гормоны железы — производные тирозина. Тироксин (T4) и трийодтиронин (Т3) увеличивают обменные процессы, ускоряют катаболизм белков, жиров и углеводов, необходимы для нормального развития ЦНС, увеличивают ЧСС и сердечный выброс. Содержание тироксина в крови больше, чем трийодтиронина (4:1), но активность последнего в 4-10 раз больше, чем тироксина. Транспортируются белками: тироксин связан прочнее, чем трийодтиронин.
Кальцитонин (32-аминокислотный пептид) и катакальцин (21-аминокислотный пептид). Их функции антагонистичны эффектам ПТГ — гормона паращитовидной железы: кальцитонин уменьшает [Са2+] в крови, стимулирует минерализацию кости, усиливает почечную экскрецию Са2+, фосфатов и Na+ (уменьшается их реабсорбция в канальцах почки).
Относящиеся к кальцитониновому гену пептиды и (37 аминокислот) экспрессируются в ряде нейронов ЦНС и на периферии (особенно в связи с кровеносными сосудами). Их функции — участие в ноцицепции, пищевом поведении, а также в регуляции тонуса сосудов. Рецепторы к этим пептидам найдены в ЦНС, сердце, плаценте.
Йодсодержащие гормоны
Материал этого раздела см. в книге.
Тироксин
Тироксин - 3,5,3',5'‑тетрайодтиронин, образуется из пары дийодтирозинов. Тироксин — основной йодсодержащий гормон, на долю T4 приходится не менее 90% всего содержащегося в крови йода.
◊ Транспорт в крови. Не более 0,05% Т4 циркулирует в крови в свободной форме, практически весь тироксин находится в связанной с белками плазмы форме. Главный транспортный белок — тироксин–связывающий глобулин (связывает 80% Т4); на долю тироксин–связывающего преальбумина, а также альбумина приходится 20% Т4.
◊ Время циркуляции в крови (время полужизни) T4 около 7 дней, при гипертиреозе — 3-4 дня, при гипотиреозе — до 10 дней.
◊ L-форма тироксина физиологически примерно вдвое активнее рацемической (DL-тироксин), D-форма гормональной активности не имеет.
◊ Дейодирование наружного кольца тироксина, частично происходящее в щитовидной железе, осуществляется преимущественно в печени и приводит к образованию Т3.
◊ Реверсивный трийодтиронин. Дейодирование внутреннего кольца тироксина происходит в щитовидной железе, преимущественно в печени и частично в почке, в результате образуется реверсивный (обратный) Т3 (3,3',5'‑трийодтиронин, rT3 [от англ. reverse]), имеющий после рождения незначительную физиологическую активность.
Содержание rT3 в амниотической жидкости много выше rT3 крови матери. Это обстоятельство означает, что преобладающий йодсодержащий гормон плода — rT3.
Измерение rT3 в амниотической жидкости используют для диагностики возможной недостаточности функции щитовидной железы (гипотиреоз плода).
Трийодтиронин
Образуется из монойодтиронина и дийодтиронина (в ЩЖ: синтезируется около 15% циркулирующего в крови Т₃, остальной трийодтиронин образуется при монодейодировании наружного кольца тироксина, происходящем преимущественно в печени). На долю Т₃ приходится лишь 5% содержащегося в крови йода.
◊ Транспорт в крови. Не более 0,5% Т₃ циркулирует в крови в свободной форме, практически весь трийодтиронин находится в связанной с белками плазмы форме.
◊ Время циркуляции в крови (время полужизни) T4 около 1,5 дней.
◊ Физиологическая активность Т₃ примерно в 4 раза выше, чем тироксина, но время полужизни много меньше. Биологическая активность и Т₃, и Т₄ обусловлена несвязанной фракцией.
◊ Катаболизм йодтиронинов. Т₃ и Т₄ конъюгируются в печени с глюкуроновой или серной кислотами и секретируются с желчью, всасываются в кишечнике, дейодируются в почках и выделяются с мочой.
Рецепторы тиреоидных гормонов
Ядерные рецепторы тиреоидных гормонов — факторы транскрипции. Известно не менее трёх подтипов этих рецепторов: 1, 2 и .
Подтип 1 экспрессируется преимущественно в ЦНС, подтипы 2 и экспрессируется во многих органах.
Сродство Т3 к рецепторам клеток–мишеней в 10 раз выше, чем у Т4.
Нечувствительность к тиреоидным гормонам. Известно несколько десятков дефектов генов, приводящих на фоне различной выраженности гипертиреоза к развитию различных синдромов (например, к синдрому гиперактивного ребёнка, периодической тахикардии, затруднённому обучению, низкорослости, глухоте).
Функции йодсодержащих гормонов
Функции йодсодержащих гормонов многочисленны. Т3 и Т4 увеличивают интенсивность обменных процессов, ускоряют катаболизм белков, жиров и углеводов, они необходимы для нормального развития ЦНС, увеличивают ЧСС и сердечный выброс. Крайне разнообразные эффекты йодсодержащих гормонов на клетки–мишени (ими практически являются все клетки организма) объясняют увеличением синтеза белков и потребления кислорода.
◊ Синтез белков увеличивается в результате активации транскрипции в клетках–мишенях, в том числе гена гормона роста. Йодтиронины расценивают как синергисты гормона роста. При дефиците Т3 клетки гипофиза теряют способность к синтезу СТГ.
◊ Потребление кислорода возрастает в результате увеличения активности Na+,K+‑АТФазы.
◊ Печень. Йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3 повышает чувствительность клеток к эффектам адреналина (стимуляция липолиза в жировой ткани и мобилизация гликогена в печени).
◊ Мышцы. Т3 увеличивает потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы, повышает чувствительность к действию адреналина.
◊ Теплопродукция. Йодтиронины участвуют в формировании ответной реакции организма на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина.
◊ Гиперйодтиронинемия. Очень высокие концентрации йодтиронинов тормозят синтез белков и стимулируют катаболические процессы, что приводит к развитию отрицательного азотистого баланса.
◊ Оценка функций щитовидной железы
Радиоиммунологический анализ позволяет прямо измерять содержание Т3, Т4, тиреотропного гормона (ТТГ).
Поглощение гормонов смолами — непрямой метод определения связывающих гормоны белков.
Индекс свободного тироксина — оценка свободного Т4.
Тест стимуляции ТТГ тиреолиберином определяет секрецию в кровь тиреотропина в ответ на внутривенное введение тиреолиберина.
Тесты выявления АТ к рецепторам ТТГ выявляют гетерогенную группу Ig, связывающихся с рецепторами ТТГ эндокринных клеток щитовидной железы и изменяющих её функциональную активность.
Сканирование щитовидной железы при помощи изотопов технеция (99mТс) позволяет выявить области пониженного накопления радионуклида (холодные узлы), обнаружить эктопические очаги щитовидной железы или дефект паренхимы органа. 99mТс накапливается только в щитовидной железе, период полувыведения составляет всего 6 часов.
Исследование поглощения радиоактивного йода при помощи йода-123 (123I) и йода-131 (131I).
Содержание йода в питьевой воде. Проводится йодирование воды на водопроводных станциях.
Пищевая соль. В России запрещено выпускать нейодированную пищевую соль.
◊ Тиреоидный статус определяет эндокринную функцию щитовидной железы. Эутиреоидия — отсутствие отклонений. Заболевание щитовидной железы можно предположить при появлении симптомов недостаточности эндокринной функции (гипотиреоз), избыточных эффектов тиреоидных гормонов (гипертиреоз) либо при очаговом или диффузном увеличении щитовидной железы (зоб).
Избыточная секреция тиреоидных гормонов.
Повышает основной обмен на 60–100% и уменьшает массу тела.
Ускоряет потребление глюкозы клетками, усиливает гликолиз и глюконеогенез, увеличивает скорость абсорбции углеводов из кишечника и стимулирует выделение инсулина.
Понижает концентрацию холестерола, фосфолипидов и триглицеридов в плазме, увеличивает концентрацию свободных жирных кислот.
Повышает тканевой метаболизм, ускоряет потребление O2 и выделение продуктов обмена, что значительно усиливает кровоток. В соответствии с увеличением кровотока растёт сердечный выброс.
Значительно повышает возбудимость и увеличивает ЧСС, но подавляет силу сокращений сердца вследствие усиленного распада белков и экспрессии тяжёлой цепи с более высокой активностью АТФазы миозина.
Увеличивает потребление O2 и образование CO2. Эти эффекты активируют все механизмы, повышающие ЧД и ГД.
Стимулирует секрецию желудочного сока и моторику ЖКТ.
Повышает возбудимость ЦНС, увеличивает реактивность синапсов в спинном мозге, что проявляется лёгким тремором кончиков пальцев с частотой 10–15 в секунду. Увеличивает чувствительность адренорецепторов к норадреналину.
Увеличивает секрецию эндокринных желез и повышает потребность тканей в гормонах. Так, увеличение секреции T4 усиливает метаболизм глюкозы во всех тканях организма, соответственно возрастает необходимость в инсулине.
Стимулирует остеогенез.
Увеличивает скорость инактивации ГЛК в печени, что — по механизму обратной связи — стимулирует секрецию АКТГ в передней доле гипофиза и ГЛК в коре надпочечников.
Недостаточная секреция тиреоидных гормонов.
Снижает на 50% основной обмен, увеличивает МТ.
Повышает концентрацию в плазме холестерола, фосфолипидов и триглицеридов.
Уменьшает сердечный выброс.
Вызывает сонливость.
У мужчин приводит к потере либидо. У женщин нарушается регулярность менструального цикла, возможны интенсивные и частые менструальные кровотечения.
Таблица. Метаболические эффекты йодсодержащих гормонов ЩЖ
-
Гипотиреоз
Гипертиреоз
Основной обмен
Масса тела
Обмен углеводов
норма
Глюконеогенез
Гликогенолиз
Глюкоза плазмы
норма
Обмен белка
Синтез
Протеолиз
Обмен липидов
Липогенез
Липолиз
Холестерол крови
Термогенез
Симпатическая нервная система
норма
норма
Катехоламины
-Адренергические рецепторы
норма