

[Rukavishnikov_A.I.]_Azbuka_raka(BookFi.org)
.pdfФакторы роста и их рецепторы раковой клетки, G-белки, связанные с мембраной клетки, белки-передатчики сигнала в цитоплазме, белки транскрип-
ции в ядре клетки – это новые мишени для создания избирательно действую-
щих средств и лекарств против раковых клеток. Большой вклад в это внесли учѐные нашей страны и их материалы мы здесь цитируем – Н.Е. Кушлинский,
Е.С. Герштейн (1996), С.А. Тюляндин (1999), Н.Е. Кушлинский (1999), Д.А.
Носов (2001).
1. В раковой клетке чрезмерная экспрессия рецептора факторов роста.
Например, рецептор эпидермального фактора роста – EGFR. Он продукт гена c-erbB1. За счет амплификации гена, т.е. избытка его иРНК, синтез рецеп-
тора чрезмерен.
Для подавления чрезмерной экспрессии рецептора, можно подавлять этот ген разными способами, например, интерферирующей РНК (РНКи) к иРНК этого гена. Можно избирательно связать белок-рецептор моноклональным ан-
тителом или синтезировать по его пространственной структуре избирательно действующее химическое соединение. Молекула этого соединения будет ком-
плементарна активным участкам молекулы-рецептора. В результате будет пре-
кращена передача сигнала к делению в раковой клетке, что затем может вести к регрессу рака.
2. Тирозинкиназа С. Она составляет цитоплазматическую часть белка-
рецептора для многих факторов роста – эпидермальный фактор роста (EGF) и TGFs-альфа, фактор роста эндотелия сосудов (VEGF-1) и другие. Присоединяя фосфатную группу, этот фермент становится активным и затем фосфорилирует молекулы-передатчики сигналов. Для подавления тирозинкиназы С уже синте-
зированы химические соединения, которые ингибируют фосфорилизацию еѐ,
что исключает сигнал от этого фермента к молекулам-передатчикам сигналов в раковой клетке.
3. Эпидермальный фактор роста (EGF) и трансформирующий фактор рос-
та (TGFs-aльфа). Они имеют общий рецептор на раковой клетке. Эти молекулы-
лиганды при связывании с рецептором на раковой клетке, вызывают еѐ деле-
90

ние. В раковой клетке их синтез чрезмерен. Здесь могут быть два воздействия:
подавление синтеза и секреции факторов роста в раковой клетке и блокада свя-
зывания его с молекулой-рецептором на раковой клетке. Для этого создаются химические соединения. Но проще создать интерферирующую РНК к иРНК ге-
на фактора роста и/или то же к гену белка-рецептора этого фактора роста.
4. Белок гена семейства ras. Из-за мутации в этом гене белок Ras не спо-
собен терять фосфатную группу, поэтому постоянно активен, стимулируя рако-
вую клетку к делению.
Белок гена ras для выполнения своего действия должен прежде приобре-
сти необходимую для него пространственную структуру и прикрепиться к внутренней поверхности мембраны раковой клетки. Для этого ему требуется фермент – фарнезилтрансфераза. Без него белок гена ras не может присоеди-
нять к себе фосфатные группы, а значит передавать сигналы от белка-рецептора к ядру раковой клетки. Для подавления фермента создаются ингибиторы его,
что будет тормозить пролиферацию раковой клетки.
5. Раковая клетка-мишень для лекарств и других средств. На раковой клетке имеются белки-рецепторы к эпидермальному фактору роста, к эмбрио-
нальному белку альфа-фетопротеину. На этой основе можно создать гибридный белок из этого фактора роста или альфа-фетопротеина, к нему химически при-
соединить молекулу цитотоксического препарата. При этом молекула белка служит средством адресной доставки, т.е. только в раковые клетки, не затраги-
вая здоровые клетки, и так уничтожать их.
Можно создать моноклональное антитело к р-гликопротеину – он на ра-
ковой клетке любого типа. Его химически соединить с цитотоксическим препа-
ратом и также адресно доставить в раковые клетки. Побочного эффекта у паци-
ента может не быть совсем или будет минимальным, так как на раковой клетке экспрессия р-гликопротеина намного больше, чем на нормальной клетке.
6. Внедрение нормальной копии гена белка р53 в раковые клетки.
При наличии мутаций в гене этого белка, в раковые клетки можно вво-
дить с помощью вирусного вектора, в том числе вируса Т4, нормальную копию
91

гена р53. Это восстановит апоптоз в раковых клетках у пациента, что и унич-
тожит их.
Наличие мутаций в гене р53 – ценный маркер для ранней диагностики ра-
ковых клеток в организме пациента по анализу образца плазмы крови от него.
Итак, лекарства стандартной химиотерапии направлены для уничтожения всех раковых клеток в организме пациента. Но это не удается достичь, так как сами лекарства не могут отличить раковую клетку от нормальной клетки. Ми-
шенью для них является ДНК раковых клеток, она же оказывается мишенью в той же степени и для нормальных клеток.
Лекарства, действующие на факторы роста и молекулы – передатчики сигналов в раковых клетках, «больше подавляют пролиферацию раковых кле-
ток, но не убивают их». Но в отличие от химиотерапии, их действие избира-
тельное – на раковые клетки. Это новый метод, для которого необходимо выяв-
лять в раковой клетке от конкретного пациента генетические и биохимические
«отклонения» (С.А. Тюляндин, 1999).
В разделе 3.2. мы рассматривали GPCR и G-белок в передаче сигналов в нормальной клетке.
Т. Кенакин (T. Kenakin, 2006) отмечает, что GPCR-рецепторы имеют еще одно свойство – «конститутивную активность». То есть «иногда они активиру-
ют G-белки без всякой видимой причины, т.е. в отсутствие лиганда».
Учѐный подчеркивает, что на раковых клетках «необычно много консти-
тутивных рецепторов» и предполагает, что с этим может быть связана «их спо-
собность к бесконтрольному делению». Такой «сбой в поведении раковой клет-
ки от конститутивных рецепторов не удаѐтся устранить никакими известными сегодня лекарственными средствами» (Рис. 2).
92
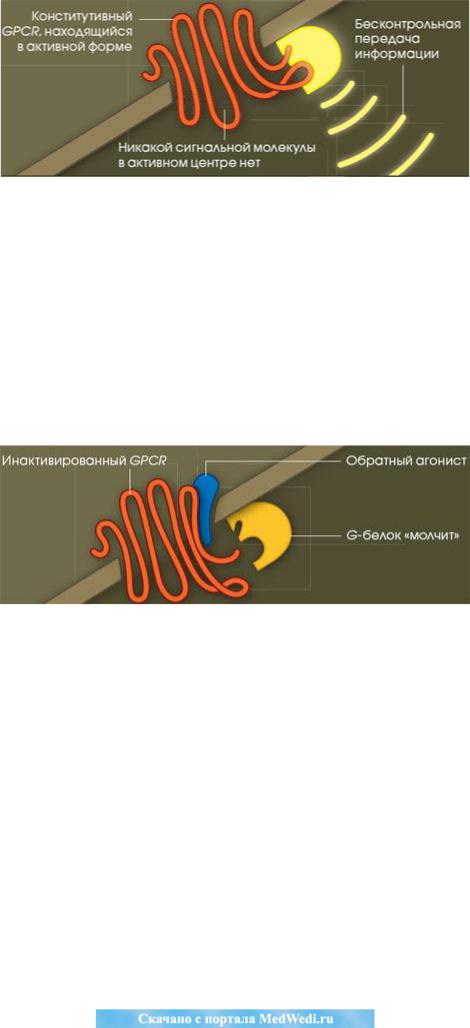
Рис. 2. Конститутивный GPCR-рецептор в активной форме в отсутствие лиганда (рис. и цит. по: Т. Кенакин, 2006).
Для этого необходимы лекарства «совершенно иного действия, которые фиксировали бы конститутивные GPCR-рецепторы в неактивной конформа-
ции».
Автор предлагает для достижения этой цели ингибитор – обратный инги-
битор или агонист, который связывается с этим рецептором и инактивирует его,
устраняя его контакт с G-белком, и прерывая этим сигнал (Рис. 3).
Рис. 3. Инактивация GPCR-рецептора и прерывание сигнала присоедине-
нием обратного агониста к рецептору (рис. и цит. по: Т. Кенакин, 2006).
Проф. Джон Дэйви, – факультет биологии Варвикского университета и компания «Септеген» (Англия) в 2003 г. разработали технологию быстрого ис-
пытания лекарств для лечения различных болезней, в том числе и рака.
Мишени этих лекарств – рецепторы, связанные с G-белками (GPCR), ко-
торые обеспечивают передачу сигналов между клетками в организме. Дефекты этих белков ведут ко многим болезням и к возникновению раковой клетки. Это причина того, что связанные с ними рецепторы стали той мишенью, на которую должны воздействовать новые лекарства.
93

Секвенирование генома человека дало специалистам информацию о функции нескольких сотен рецепторов, многие из которых могут стать мише-
нями лекарственного воздействия. Однако, говорит проф. Джон Дэйви, «знать,
что эти мишени или цели существуют – это одно, а способность воздействовать на них – совсем другое».
Проф. Джон Дэйви разработал новый метод, который позволяет выявить возможное воздействие лекарств на эти рецепторы.
По технологии GPCR человека помещают в живую клетку простых дрожжей и наблюдают за действием лекарства. По мнению автора, новая тех-
нология позволяет «определять мельчайшие различия между рецепторами раз-
ных людей, что дает возможность выявить лекарства, наиболее эффективные для каждого пациента».
94

Глава 5. Клеточный цикл. Молекулы-регуляторы клеточного цикла
открывают пути к диагностике и уничтожению раковых клеток
В организме взрослого человека 5∙1013 (В.Н.Сойфер, 1998) или 5∙1014 (В.
Тарантул, 2003) клеток. Каждая клетка любого типа – это часть своей ткани и организма в целом.
Раковая клетка в организме человека – это уже не часть ткани и своего организма, а самостоятельная клетка, отделившаяся от них. Это клетка-
организм.
Деление клетки – это основное свойство и признак того, что она живая. В
раковой клетке свойство деления нарушено, и она делится без конца.
Если в организме нормальная клетка того или иного типа делится для су-
ществования организма, то размножение раковой клетки – для его уничто-
жения.
То, что клетки размножаются путем деления, известно более 100 лет. Но почему клетка делится, оставалось не ясно до конца 70-х годов XX века.
Каждая клетка после деления делает выбор: либо она начинает синтез ДНК и в таком случае будет вновь делиться, либо она избирает путь к диффе-
ренцированной клетке и это означает, что она уже больше никогда не разделит-
ся. Молекулярные причины, регулирующие этот «выбор» долго оставались не выясненными, а это важно для понимания превращения ее в раковую клетку.
Период жизни клетки от одного деления до другого или от деления до еѐ смерти – это клеточный цикл. Он состоит из интерфазы – вне деления, и самого деления клетки. Интерфаза – период, во время которого клетка растѐт и удваи-
вает все свои компоненты. Митоз – это процесс, в результате которого из одной клетки образуется две. Он из двух стадий: митоз – деление клеточного ядра, и
цитокинез – деление клетки на две дочерние клетки.
В интерфазе различают три этапа или фазы: G1, S, G2, а в митозе четыре:
профаза, метафаза, анафаза и телофаза.
Этапы интерфазы
95

1.G1 фаза (от англ. ―gap 1‖, то есть интервал 1) – рост клетки и удвоение всех еѐ компонентов.
2.S фаза (от ―synthesis‖) – синтез ДНК путем репликации, формирование копий каждой хромосомы; удвоение центросомы.
3.G2 фаза (―gap 2‖) – подготовка к митозу, продолжение роста клетки,
синтез необходимых белков.
Этапы или фазы митоза
M фаза (от. ―mitosis‖) – деление клетки на две дочерние. В митозе четыре фазы, им заканчивается клеточный цикл.
Профаза. Каждая хромосома – это пара хроматид, соединѐнных в месте центромеры. Центриоли расходятся к противоположным полюсам клетки. От каждой центриоли в виде лучей расходятся микротрубочки, многие из которых становятся фибриллами митотического веретена. В каждой хроматиде в обла-
сти центромеры имеется богатый белком участок – кинетохор – важный эле-
мент веретена. Кинетохоры в роли сцепления, которое позволяет взаимодей-
ствовать фибриллам веретена с хромосомами: как только к кинетохору присое-
диняется микротрубочка, хромосома начинает двигаться. Ядерная мембрана распадается и образуется веретено деления.
Метафаза. Хроматиды прикрепляются к фибриллам веретена кинетохо-
рами. Оказавшись связанными с обеими центросомами, хроматиды движутся к экватору веретена до тех пор, пока их центромеры не выстроятся по экватору веретена перпендикулярно его оси. Это позволяет хроматидам беспрепятствен-
но двигаться к соответствующим полюсам. Характерное для метафазы разме-
щение хромосом очень важно для сегрегации хромосом, т.е. расхождения сест-
ринских хроматид. Если отдельная хромосома «замешкается» в своѐм движе-
нии к экватору веретена, задерживается обычно и начало анафазы. Метафаза завершается разделением сестринских хроматид.
Анафаза. Каждая центромера расщепляется на две, нити веретена оття-
гивают дочерние центромеры к противоположным полюсам. Центромеры тянут
96

за собой отделившиеся одна от другой хроматиды, которые теперь становятся независимыми хромосомами.
Телофаза. Хромосомы достигают полюсов клетки, деспирализуются, и их уже нельзя четко различить. Нити веретена разрушаются, а центриоли репли-
цируются. Вокруг хромосом на каждом из полюсов образуется ядерная оболоч-
ка. За телофазой может сразу следовать цитокинез – разделение всей клетки на две (Рис. 1). Каждая из дочерних клеток имеет идентичный набор хромосом.
После деления клеточный цикл завершается, и клетка переходит в фазу G1.
Длительность цикла определяется типом клетки. Обычно она составляет от 10 до 30 часов. Клетки в фазе G1 не всегда делятся, они могут выйти из цик-
ла и перейти на фазу покоя – G0 фаза.
Лишь после 1970-х годов было установлено, что прохождение клетки по всем фазам клеточного цикла строго регулируется. При движении по циклу в них появляются и исчезают, активируются и ингибируются регуляторные мо-
лекулы, которые обеспечивают: 1) прохождение клетки по определенной фазе клеточного цикла и 2) переход из одной фазы в другую. Причѐм прохождение по каждой фазе, а также переход из одной фазы в другую контролируется ра-
зличными ключевыми молекулами. Они регулируют клеточный цикл в орга-
низмах всех эукариотов – дрожжей, животных и человека.
97
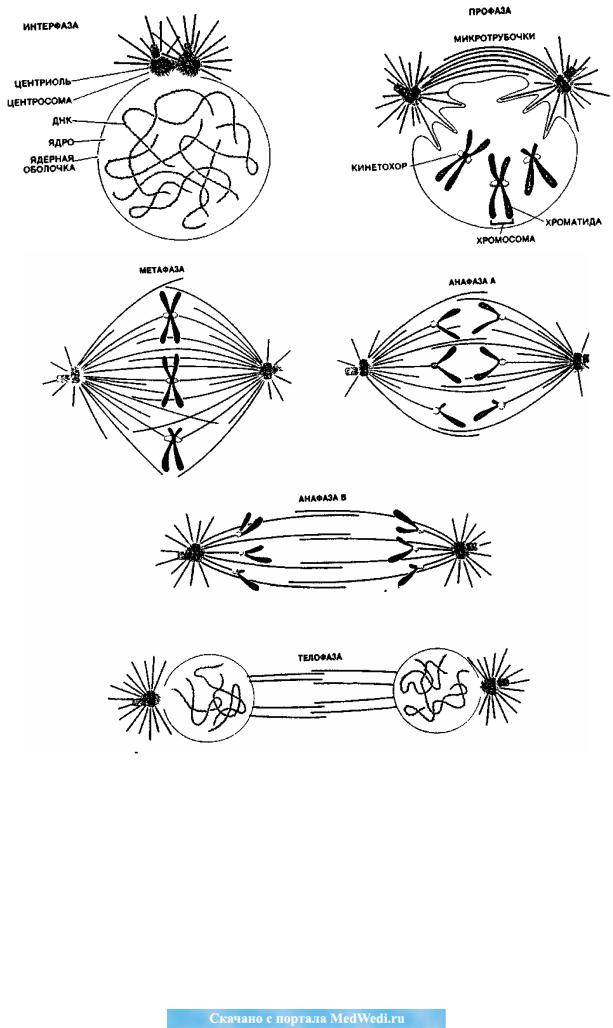
Рис. 1. Схемы последовательных фаз митоза в эукариотической клетке
(цит. и рис. по: Дж.Р. Макинтош, К.Л. Макдоланд (1989) с изменениями).
Теперь мы кратко изложим, что это за ключевые молекулы, и что они де-
лают в клетке. Ясно, что мы будем называть учѐных, которые открыли эти мо-
лекулы, и вклад каждого из них в эти знания. Пока мы скажем, что результаты их исследований очень важны для понимания любой болезни, а в онкологии
98

могут произвести настоящую революцию как в диагностике рака, так и в изле-
чении от него.
Л. Хартуэлл (L. Hartwell) из США в 1970-1971 гг. впервые для изучения клеточного цикла применил генетические методы. Он первым для этого ис-
пользовал клетки пекарских дрожжей. Дрожжевая клетка – удобная модель для этого, так как: умеет размножаться, отвечать на сигналы внешней среды, осу-
ществлять «ремонт» генов при мутациях от влияния среды. Кроме того, клетки дрожжей имеют гены, родственные всем основным генам человека. Идея учѐ-
ного: изучение клеточного цикла и его регуляции в клетках дрожжей может помочь понять это в нормальной клетке человека, а затем и в раковой клетке.
В серии опытов Л. Хартуэлл выделил клетки дрожжей, в которых гены,
управляющие циклом, были изменены. Так он выделил более 100 генов, участ-
вующих в управлении клеточным циклом – гены сдс (от англ. – гены цикла де-
ления клетки). Наиболее важным из них оказался ген сдс28 – «стартовый». Он управляет первым шагом в прохождении клеточного цикла через фазу G1.
Он изучал чувствительность клеток дрожжей к радиации. Оказалось, что ответом клетки на это – остановка клеточного цикла. Учѐный смог выделить гены, которые в этих опытах тормозили клеточный цикл клетки. Их оказалось много, он дал им имя – rad.
Белок таких генов «чувствует», что в ДНК клеток от облучения появи-
лись «разрывы» или неправильно спаренные основания. Другие белки этих ге-
нов проверяют состояние ДНК. Для этого они останавливают клеточный цикл,
а ферменты репарации ДНК производят еѐ «ремонт».
На основе этих результатов Хартуэлл ввел понятие «контрольной точки».
На этом этапе проверяется идентичность ДНК: если она повреждена, то цикл останавливается, чтобы дать время для исправления ДНК прежде, чем клетка перейдѐт в следующую фазу. Если исправление ДНК невозможно, клетка полу-
чает сигнал на апоптоз.
Теперь такая проверка осуществляется в нескольких точках клеточного цикла, получивших название сверочных точек – чекпоинтов (checkpoint). Эти
99