

физиол мышц1
.pdf21
мышцы в момент сокращения происходит в результате активного скольжения актиновых нитей относительно миозиновых нитей. При этом саркомер спосо-
бен укоротиться на 20-30% своей длины. Основные положения этой теории:
а) ионы Ca2+ запускают механизм мышечного сокращения; б) за счет ионов Ca2+
происходит скольжение тонких актиновых нитей по отношению к миозиновым.
4. Взаимодействие ионов Са2+ с тропонином, освобождение активных центров на актиновых филаментах.
5. Взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги.
6. Скольжение нитей актина и миозина относительно друг друга, уменьше-
ние размера саркомера, развитие напряжения или укорочение мышечного во-
локна.
В покое взаимодействие тонких и толстых нитей невозможно, т.к. миозин-
связывающие участки актина заблокированы тропомиозином (рис. 10).
При высокой концентрации Ca2+ эти ионы связываются с тропонином C и
вызывают конформационные изменения тропомиозина, приводящие к разбло-
кированию миозин-связывающих участков. От концентрации Ca2+ в цитоплазме зависит количество занятых Ca2+-связывающих участков тропонина, что в свою очередь, определяет, какое количество участков актина доступно для взаимо-
действия с поперечными мостиками.
Рис. 10. Ca2+–зависимый механизм регуляции взаимодействия актина с миозином

22
Головки миозина, несущие продукты гидролиза АТФ (АДФ + Фн), присо-
единяются к тонкой нити и изменяют свою конформацию, создавая тянущее усилие: — тонкие нити начинают скользить между толстыми. За счёт шарнир-
ного участка в области шейки миозина происходит гребковое движение, про-
двигающее тонкую нить к центру сaркомера. В результате происходит сколь-
жение тонких нитей относительно толстых. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. Последующий гидролиз АТФ восстанавливает конформированную молекулу миозина, гото-
вую вступить в новый цикл.
Каждый цикл взаимодействия тонких и толстых нитей имеет несколько стадий (рис. 11). Последовательность событий, начиная от связывания попе-
речного мостика с тонким филаментом и до момента, когда система готова к повторению процесса называется рабочим циклом поперечных мостиков.
Рис. 11. Цикл взаимодействия тонких и толстых нитей: А- состояние покоя; Б- прикрепление головки миозина к актину, поворот головки миозина; В- высвобождение АДФ и Фн, развитие тянущего усилия, присоединение новой молекулы АТФ, что приводит к отделению головки миозина от молекулы актина; Г-гидролиз АТФ возвращает молекулу миозина в исходное положение.
23
Каждый цикл состоит из 4 стадий:
1)Прикрепление поперечного мостика к актиновому филаменту (образование актомиозиновых мостиков увеличивает АТФазную активность головок мио-
зиновых нитей и освобождение энергии АТФ)
2)Гребковое движение (деформация актомиозиновых мостиков в шарнирных участках, что вызывает смещение актиновой нити вдоль миозиновой на 1 %
длины)
3)Отсоединение поперечных мостиков
4)Получение энергии (при присоединении АТФ к головке миозина и ее гидро-
лизе) - головка присоединяется в новом месте Таким образом, в цикле поперечных мостиков АТФ выполняет две разные ро-
ли: 1) его гидролиз поставляет энергию для движения поперечного мостика; 2)
его связывание (но не гидролиз) с миозином сопровождается отделением по-
следнего от актина и создает возможность повторения цикла поперечных мо-
стиков.
Расслабление мышц. В настоящее время расслабление считается актив-
ным процессом, причем расход энергии больше, чем при сокращении. Источ-
ник сокращения - АТФ, расщепляющийся при сокращении.
Доказательства:
-появление мышечных контрактур при утомлении (т. е. при деятельном сокра-
щении);
-явления трупного окоченения (отсутствие энергии).
Для расслабления миоцита необходимы два главных условия:
1)Наличие достаточного уровня АТФ
2)Низкая концентрация Са 2+ - ниже до 10 -7 (создается активацией Са 2+-
насоса, перемещение Са 2+- в цистерны саркоплазматического ретикулума,
снижение концентрации Са 2+- в саркоплазме, восстановление блокады тро-
помиозином миозинсвязывающих участков актиновых нитей).
Контрольные вопросы
1. Что такое электромеханическое преобразование в мышечном волокне?
24
2.Какие основные положения теории «скользящих нитей» О. Хаксли?
3.Из каких этапов состоит сокращение мышцы?
4.Из каких стадий состоит цикл взаимодействия актина с миозином?
5.При каких условиях происходит расслабление мышцы?
1.6. Химические и тепловые процессы при сокращении мышц
Мышечное сокращение требует значительных энергетических затрат.
Универсальным непосредственным носителем энергии в клетках является АТФ.
Энергия АТФ в скелетной мышце используется для трех процессов:
1) работы натрий-калиевого насоса, обеспечивающего поддержание постоян-
ства градиента концентрации этих ионов по обе стороны мембраны; 2) процесса скольжения актиновых и миозиновых нитей, приводящих к укорочению мио-
фибрилл; 3) работы кальциевого насоса, активируемого при расслаблении волокна.
Расход энергии: на процесс сокращения - 70 %, на процесс расслабления -
15 %, на работу насоса- 5 %, на синтез -10 %.
На одно рабочее движение 1 мостика тратится 1 молекула АТФ. В мышеч-
ном волокне концентрация АТФ равна 4 ммоль/л. Такого запаса энергии доста-
точно для поддержания сокращения не более 1–2 сек. Поэтому должен посто-
янно происходить ресинтез АТФ.
1. Анаэробные пути ресинтеза АТФ:
А. фосфагенная, или креатинфосфатная система
Первым источником для восстановления АТФ является использование кре-
атинфосфата — вещества, имеющего высокоэнергетические фосфатные связи,
подобные связям АТФ. Однако количество креатинфосфата в мышечных во-
локнах невелико, всего на 1/5 больше, чем АТФ. Общих запасов энергии АТФ и креатинфосфата в мышечном волокне достаточно для развития макси-
мального мышечного сокращения лишь в течение 5–8 сек.
25
Б. лактацидная система
Расщепление гликогена до пировиноградной и молочной кислот сопро-
вождается выделением энергии, которая идёт на превращение АДФ в АТФ.
Вновь синтезированный АТФ может использоваться или непосредственно для мышечного сокращения, или в процессе восстановления запасов креатинфос-
фата. Гликолитические реакции могут происходить в отсутствие кислорода, и
мышца может сокращаться десятки секунд без снабжения кислородом. Однако большое количество промежуточных продуктов гликолитического обмена (за минуту образуется 20 мМ молочной кислоты), накапливаемых в мышечных во-
локнах, не позволяет гликолизу поддерживать максимальное сокращение более одной минуты. Предельное время для такого рода работы составляет около 30
с, после чего возникает накопление молочной кислоты, т. е. метаболический ацидоз, и развивается утомление.
2. Аэробный путь ресинтеза АТФ, или окислительное фосфорилиро-
вание. Третьим источником энергии является окислительный метаболизм. Бо-
лее 95% энергии, используемой мышцей в ходе продолжительных, напряжён-
ных сокращений, поступает именно из этого источника. В качестве субстратов используются в основном углеводы и жиры, роль белков ничтожна. При уме-
ренной мышечной активности АТФ образуется преимущественно путем окис-
лительного фосфорилирования, и в течение первых 5—10 мин главным ресур-
сом для этого служит гликоген. В последующие 30 мин доминирующими ста-
новятся источники энергии, доставляемые кровью, причем глюкоза и жирные кислоты участвуют примерно в одинаковой мере. На более поздних этапах со-
кращения преобладает утилизация жирных кислот, а глюкоза расходуется меньше (рис.12).
Энергетические системы, используемые при разных видах спорта пред-
ставлены в приложении 2, таблице 3.

26
Рис. 12. Источники энергии для работы мышцы. В разные периоды работы мышцы ее энергетические потребности покрываются за счет различных процессов
Жировой запас энергии значительно превышает углеводный (табл.3).
|
|
|
Таблица 3 |
|
Запасы "топлива" и энергии в организме |
|
|
||
|
|
|
|
|
источник энергии |
г |
|
ккал |
|
|
|
|
|
|
углеводы: |
|
|
|
|
|
|
|
|
|
гликоген печени |
110 |
|
0, 451 |
|
|
|
|
|
|
мышечный гликоген |
250 |
|
1,025 |
|
|
|
|
|
|
глюкоза в жидкостях организма |
15 |
|
0,062 |
|
|
|
|
|
|
всего: |
375 |
|
1,538 |
|
|
|
|
|
|
жиры: |
|
|
|
|
|
|
|
|
|
подкожный |
7,8 |
|
70,980 |
|
|
|
|
|
|
внутримышечный |
0,161 |
|
1,465 |
|
|
|
|
|
|
всего |
7,961 |
|
72,445 |
|
|
|
|
|
|
Примечание: оценка произведена на основании средней массы тела 65 кг с содержанием жи-
ра 12 %
Белки как энергетический субстрат используются крайне редко – только после того, как будут исчерпаны имеющиеся запасы углеводов и жиров. По-
требность спортсменов, тренирующихся в различных видах спорта в белках,

27
жирах, углеводах, витаминах и минеральных веществах представлены в табли-
цах 5, 6, 7, 8 приложения 4.
По окончании мышечной работы запасы богатых энергией соединений — креатинфосфата и гликогена — в мышце снижены и для возвращения исходно-
го состояния мышечного волокна необходимо, чтобы они пополнились.Для восстановления запасов обоих соединений нужна энергия, поэтому мышца, уже будучи в состоянии покоя, продолжает некоторое время усиленно потреблять кислород; наглядный тому пример — глубокое и учащенное дыхание сразу по-
сле интенсивной физической работы. Благодаря усиленному потреблению кис-
лорода в период после мышечной работы, покрывается так называемый кисло-
родный долг (рис. 13). Таким образом, даже на ранних стадиях тяжелой физи-
ческой работы способность человека к синтезу энергии аэробным путем ча-
стично снижается, что вызвано двумя причинами: 1) кислородным долгом; 2)
истощением запасов гликогена в мышцах.
Рис. 13. Кислородная задолженность
В норме в организме имеется примерно 2 л кислорода, который может быть использован для аэробного метаболизма даже без вдыхания новых порций кислорода. В этот запас входят: 1) 0,5 л, находящихся в воздухе легких; 2) 0,25
л растворенного в жидкостях организма; 3) 1л, связанный с гемоглобином; 4) 0,3 л в составе миоглобина. При тяжелой физической работе почти весь запас
28
кислорода используется для аэробного метаболизма в течение примерно 1
мин.Кислородная задолженность погашается после работы за счет автоматиче-
ской мобилизации дыхания и кровообращения. Раннюю часть кислородного долга (примерно 3,5 л) называют алактацидным (не связанным с молочной кис-
лотой), а позднюю (примерно 8 л) – лактацидным (связанным с удалением мо-
лочной кислоты). Если работа продолжается, то возникает утомление.
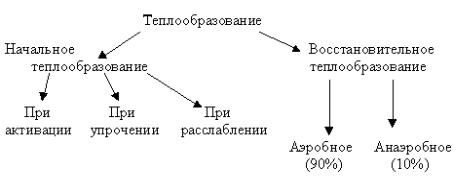
Теплообразование при мышечном сокращении
В работающей мышце доля энергии, потраченной на выполнение механи-
ческой работы приблизительно в пять раз меньше энергии, высвобождаемой в виде теплоты. По характеру своего происхождения и времени развития тепло-
образование подразделяют на две основные фазы: фазу начального теплообра-
зования и фазу восстановительного теплообразования (схема 1).
Схема 1. Различные фазы теплообразования в мышцах млекопитающих.
Фаза начального теплообразования примерно в 1000 раз короче фазы вос-
становительного теплообразования. Она начинается с момента возбуждения мышцы и продолжается в течение всего сокращения, включая фазу расслабле-
ния. А. Хиллом было установлено, что начальное теплообразование можно раз-
делить на несколько компонентов:
1. Теплота активации — быстрое выделение тепла на ранних этапах мышеч-
ного сокращения, когда отсутствуют видимые признаки укорочения или разви-
тия напряжения. Теплообразование на этой стадии обусловлено выходом ионов Са2+ из триад и соединением их с тропонином.

29
2.Теплота укорочения — выделение тепла при совершении работы, если речь идет не об изометрическом режиме. При этом, чем больше совершается механической работы, тем больше выделяется тепла.
3.Теплота расслабления — выделение тепла упругими элементами мышцы при расслаблении. При этом выделение тепла не связано непосредственно с процессами метаболизма.
Впроцессе образования тепловой энергии в мышце выделяют 2 фазы (рис. 14).
Рис. 14.Тепловые процессы в мышцах
Первая фаза начального теплообразования начинается с момента воз-
буждения мышцы, продолжается в течение всего сокращения, включая и фазу расслабления. В эту фазу, особенно в латентный период величина теплопро-
дукции в единицу времени наивысшая (до 3,5 условных единиц) и наименьшая в начальный период расслабления мышцы. Тепловая энергия образуется в ре-
зультате химических процессов расщепления АТФ, обеспечивающих возбуж-
дение, сокращение и расслабление мышцы.
Вторая фаза теплообразования длится несколько минут после расслабле-
ния мышцы и называется фазой отставленного теплообразования или восстано-
вительного. Она обусловлена процессами, обеспечивающими ресинтез АТФ.
Главную роль в ресинтезе АТФ и восстановительном теплообразовании играют процессы гликолиза и окислительного фосфорилирования. Примерно 90% вос-
30
становительного тепла образуется за счет окислительных процессов, и лишь
10% обеспечивается анаэробными процессами. В первую фазу выделяется око-
ло 40%, а во вторую - около 60% всей тепловой энергии, образовавшейся в мышце.
Как уже указывалось, не вся энергия мышечного сокращения направляет-
ся на выполнение механической работы. Значительная часть ее рассеивается в виде тепла. Коэффициент полезного действия мышечной работы (КПД) – это отношение величины внешней механической работы (W) к общему количеству выделенной в виде тепла энергии (Е):
КПД= W/Е 100%
Наиболее высокий показатель КПД изолированной мышцы наблюдается при внешней нагрузке, составляющей 50 % от ее максимального значения, и
при скорости укорочения мышцы в пределах 30 % ее максимума. В таких усло-
виях КПД составляет 20-30 %. Эти результаты, полученные на изолированных мышцах, близки к результатам исследований на человеке. Во время мышечной работы показатели КПД составляют 15-30 % в зависимости от характера рабо-
ты (интенсивность, срочность, факторы внешней среды и т.п.), соотношения динамического и статического напряжения, набора мышечных групп, реализу-
ющих двигательную задачу.
Контрольные вопросы
1.На какие процессы используется энергия АТФ в скелетной мышце?
2.Какие существуют пути ресинтеза АТФ?
3.Какие существуют анаэробные пути ресинтеза АТФ?
4.В чем отличие аэробного пути ресинтеза АТФ от анаэробных?
5.Что такое кислородная задолженность?
6.Какие виды теплообразования происходят в мышце при сокращении?
7.Чем обусловлено начальное и отставленное теплообразование?
8.Как определяется и чему равен КПД мышцы?