

физиол мышц1
.pdf11
Тропонин и тропомиозин играют важную роль в механизмах взаимодей-
ствия актина и миозина.
Контрольные вопросы
1.Что входит в Т-систему мышечного волокна?
2.Что такое саркомер?
3.Из чего состоят А-диски и I-диски?
4.Каково строение молекулы миозина?
5.Чем представлены актиновые филаменты?
1.2 Двигательные единицы
Двигательная единица (ДЕ) - простейшая структурнофункциональная единица двигательной системы, представляющая собой мотонейрон вместе с совокупностью мышечных волокон скелетной мышцы, иннервируемых этим мотонейроном (рис. 7).
Рис. 7. Двигательная единица
Термин «двигательная единица» ввел в употребление британский нейрофи-
зиолог Ч. Шеррингтон. В разных мышцах количество двигательных единиц значительно отличается. В мышцах, обеспечивающих наиболее точные и быст-
12
рые движения, двигательная единица состоит из нескольких мышечных воло-
кон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон. Так, в
латеральной прямой мышце глаза их не больше 25, в медиальной головке икро-
ножной мышцы - 1600-1700. Двигательные единицы с малым числом мышеч-
ных волокон, иннервируемых одним мотонейроном, обладают большими воз-
можностями управления сокращением. Двигательные единицы одной мышцы разные. Есть быстрые двигательные единицы, у которых скорость сокращения высокая, и медленные двигательные единицы. Последние, в свою очередь, под-
разделяются на устойчивые к утомлению и быстроутомляемые.
Порядок вовлечения двигательных единиц в работу (рекрутирование) в
обычных условиях определяется размерами мотонейронов. Первыми вовлека-
ются мотонейроны меньших размеров (медленные двигательные единицы),
развивающие небольшую силу. При увеличении уровня возбуждения (за счет увеличения частоты нервных импульсов, поступающих из двигательных цен-
тров к каждой двигательной единице) рекрутируются быстрые двигательные единицы, развивающие большую силу. Этот механизм включения двигатель-
ных единиц в соответствии с их размерами носит название "правило размера Хеннемана". Таким образом, возникает возможность точной градации напря-
жения мышцы. Важно, однако, учитывать, что в условиях обычной деятельно-
сти мышечные волокна больших двигательных единиц тренируются мало, так как развиваемые мышцами напряжения не требуют их включения; при малых напряжениях человек обходится малыми (низкопороговыми) двигательными единицами. Это происходит, например, при удержании позы, когда работают в основном единицы, устойчивые к утомлению. Мышечные волокна быстрых и медленных ДЕ также различаются между собой (таблица 1).
13
|
|
|
Таблица 1 |
Сравнительная характеристика ДЕ |
|
||
|
|
|
|
показатель |
медленные ДЕ |
|
быстрые ДЕ |
|
|
|
|
количество миоцитов |
Небольшое |
|
большое |
|
|
|
|
сила сокращения |
Небольшая |
|
большая |
|
|
|
|
утомляемость |
Низкая |
|
высокая |
|
|
|
|
Работа |
длительная маломощная |
|
кратковременная |
|
|
|
большой мощности |
|
|
|
|
тип энергообеспечения |
Аэробный |
|
анаэробный |
|
|
|
|
гладкий тетанус |
при небольшой частоте |
|
при большой частоте |
|
(<16 Гц) |
|
(>30 Гц) |
|
|
|
|
Контрольные вопросы
1.Что входит в состав двигательной единицы?
2.Какие виды двигательных единиц выделяют?
3.В чем отличия быстрых двигательных единиц от медленных?
1.3. Типы мышечных волокон
Каждая мышца — гетерогенная популяция разных типов мышечных воло-
кон. Мышечные волокна классифицируют по следующим критериям:
1)характеру сокращения:
-фазные
-тонические Фазные мышечные волокна осуществляют энергичные сокращения. Про-
извольная мускулатура человека практически полностью состоит из фазных мышечных волокон, генерирующих ПД. Тонические мышечные волокна специ-
ализированы на поддержание статического напряжения, или тонусa. Тониче-
ские мышечные волокна встречаются в наружных ушных и наружных глазных мышцах.
14
2) скорости сокращения:
-быстрые
-медленные Мышечные волокна, имеющие высокую активность АТФазы, — быстро-
сокращающиеся волокна (быстрые волокна), для медленносокращающихся во-
локон (медленные волокна) характерна низкая АТФазная активность.
3) типу окислительного обмена:
-окислительные — красные
- гликолитические — белые Окислительные, или красные мышечные волокна небольшого диаметра,
окружены массой капилляров, содержат много миоглобина. Их многочислен-
ные митохондрии имеют высокой уровень активности окислительных фермен-
тов (например, сукцинатдегидрогеназы — СДГ). Гликолитические, или белые мышечные волокна имеют больший диаметр, в саркоплазме содержится значи-
тельное количество гликогена, митохондрии немногочисленны. Для них харак-
терны низкая активность окислительных и высокая активность гликолитиче-
ских ферментов.
На практике результаты критерии разделения мышечных волокон на ти-
пы комбинируют. В настоящее время выделяют три основных типа мышечных волокон. Характеристики трех типов волокон обобщены в таблице 2.
1) Медленные фазические волокна окислительного типа (тип I). Во-
локна этого типа характеризуются большим содержанием белка миоглобина.
Мышцы, которые преимущественно состоят из волокон этого типа, за их тем-
но-красный цвет называют красными. Они выполняют очень важную функцию поддержания позы человека и животных. Предельное утомление у волокон данного типа и, следовательно, мышц наступает очень медленно, что обуслов-
лено наличием миоглобина и большого числа митохондрий. Восстановление функции после утомления происходит быстро. Нейромоторные единицы этих мышц состоят из большого числа мышечных волокон.
15
|
|
|
Таблица 2 |
Сравнительная характеристика типов мышечных волокон |
|||
|
|
|
|
показатель |
Медленные фазиче- |
Быстрые фазиче- |
Быстрые фазиче- |
|
ские волокна окис- |
ские волокна окис- |
ские волокна с |
|
лительного типа |
лительного типа |
гликолитическим |
|
(тип I) |
(тип II а). |
типом окисления |
|
|
|
(тип II б) |
главный |
окислительное |
окислительное |
гликолиз |
источник обра- |
фосфорилирование |
фосфорилирование |
|
зования АТФ |
|
|
|
количество |
много |
много |
мало |
митохондрий |
|
|
|
содержание |
высокое |
высокое |
низкое |
гемоглобина |
|
|
|
содержание |
низкое |
промежуточное |
высокое |
гликогена |
|
|
|
скорость |
медленная |
промежуточная |
быстрая |
утомления |
|
|
|
Диаметр |
малый |
промежуточный |
большой |
волокна |
|
|
|
Размер |
малый |
промежуточный |
большой |
двигательной |
|
|
|
единицы |
|
|
|
скорость |
медленная |
быстрая |
быстрая |
укорочения |
|
|
|
2) Быстрые фазические волокна окислительного типа (тип II а).
Мышцы, которые преимущественно состоят из волокон этого типа, выполняют быстрые сокращения без заметного утомления, что объясняется большим коли-
чеством митохондрий в этих волокнах и способностью образовывать АТФ пу-
тем окислительного фосфорилирования. Как правило, число волокон, входящих в состав нейромоторной единицы, в этих мышцах меньше, чем в предыдущей группе. Основное назначение мышечных волокон данного типа заключается в выполнении быстрых, энергичных движении.
3) Быстрые фазические волокна с гликолитическим типом окисления
(тип II б). Волокна данного типа характеризуются тем, что АТФ в них образу-
ется за счет гликолиза. Волокна этой группы содержат митохондрий меньше,
чем волокна предыдущей группы. Мышцы, содержащие эти волокна, развива-
16
ют быстрое и сильное сокращение, но сравнительно быстро утомляются. Мио-
глобин в данной группе мышечных волокон отсутствует, вследствие чего мыш-
цы, состоящие из волокон этого типа, называют белыми.
Контрольные вопросы
1.Как классифицируют мышечные волокна ?
2.В чем отличие фазных мышечных волокон от тонических ?
3.Как на практике используют критерии для разделения мышечных волокон на типы и какие типы выделяют ?
4.Какие характерные особенности имеются у каждого типа мышечного волок-
на ?
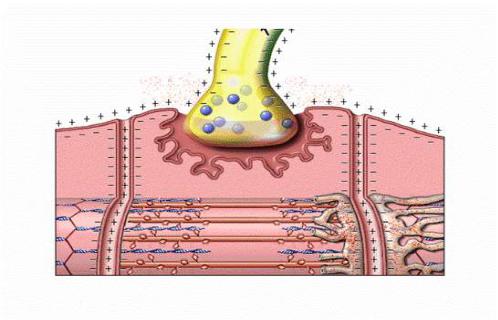
1.4. Нервно-мышечная передача
В естественных условиях (в нашем организме) возбуждение мышечного волокна (или нескольких мышечных волокон, составляющих мышцу) возникает в результате передачи возбуждения с нервного волокна на мембрану мышечно-
го в местах контакта нерва и мышцы: нервно-мышечных синапсах. Синапсы представляют собой коммуникационные структуры, которые формируются окончанием нервного волокна и прилегающей к нему мембраной мышечного волокна (рис.8).
Рис.8. Мионевральный синапс
17
На каждом мышечном волокне формируется только один синапс (за ис-
ключением примерно 2 % мышечных волокон). Как и любой синапс, нервно-
мышечное соединение состоит из трех частей: пресинаптической области,
постсинаптической области и синаптической щели. Постсинаптическую об-
ласть синапса называют двигательной концевой пластинкой.
В пресинаптической области в большом количестве присутствуют синап-
тические пузырьки, заполненные ацетилхолином (5–15 тыс. молекул в одном пузырьке) и имеющие диаметр порядка 50 нм. На постсинаптической мем-
бране — специализированной части плазмолеммы мышечного волокна — име-
ются многочисленные инвагинации, от которых на глубину 0,5–1,0 мкм отходят постсинаптические складки, чем существенно увеличивается площадь мембра-
ны. В постсинаптическую мембрану встроены Н-холинорецепторы, их концен-
трация достигает 20–30 тысяч на 1 мкм2. Через синаптическую щель проходит синаптическая базальная мембрана. Она удерживает в области синапса терми-
наль аксона, контролирует расположение холинорецепторов в виде скоплений в постсинаптической мембране. В синаптической щели также находится фермент ацетилхолинэстераза, расщепляющий ацетилхолин на холин и уксусную кисло-
ту.
Нервно-мышечная передача возбуждения состоит из нескольких этапов:
1)потенциал действия (ПД) по аксону достигает области двигательного нервного окончания
2)деполяризация мембраны нервного окончания приводит к открытию по-
тенциалозависимых Са2+-каналов и входу Са2+ в двигательное нервное оконча-
ние
3) вход Са2+ через пресинаптическую мембрану способствует высвобожде-
нию химических нейромедиаторов, находящихся в виде везикул (пузырьков) из окончания аксона
4) медиатор ацетилхолин (АХ) диффундирует через синаптическую щель и связывается со специфическими рецепторами на постсинаптической мембране
18
5)активация Н-холинорецепторов постсинаптической мембраны, при от-
крытии каналов Н-холинорецепторов возникает входящий Na–ток, что приво-
дит к деполяризации постсинаптической мембраны. Появляется потенциал концевой пластинки (ПКП), который при достижении критического уровня де-
поляризации вызывает ПД в мышечном волокне.
6) ацетилхолинэстераза (АХЭ) расщепляет ацетилхолин на составные компоненты (ацетил и холин) и действие выделившийся порции нейромедиато-
ра на постсинаптическую мембрану прекращается. Короткий период времени, в
течение которого ацетилхолин остается в синаптическом пространстве (обычно несколько миллисекунд) в норме достаточен для возбуждения мышечного во-
локна. Быстрое удаление ацетилхолина предупреждает непрерывное повторное возбуждение мышечного волокна после того, как оно восстановилось после первичного ПД. Без удаления медиатора на постсинаптической мембране раз-
вивается длительная деполяризация, которая ведет к нарушению проведения возбуждения в синапсе – синаптической депрессии.
В нервно-мышечном синапсе выделяется значительно большее количество ацетилхолина, чем требуется для запуска потенциала действия на постсинапти-
ческой мембране. Тем самым гарантируется, что каждый потенциал действия мотонейрона вызовет реакцию в иннервируемом им мышечном волокне. В
нервно-мышечном синапсе при спонтанном выходе отдельных квантов нейро-
медиатора ацетилхолина возникают миниатюрные потенциалы концевой пла-
стинки (МПКП).
Таким образом, нервно-мышечный синапс является «выгодным» местом,
куда можно воздействовать фармакологическими препаратами, изменяя чув-
ствительность рецептора, активность фермента. Эти явления будут часто встре-
чаться в практике врача: например, при отравлении токсином ботулизма – бло-
кируется высвобождение медиатора ацетилхолина (разглаживание морщин в косметической медицине), блокада холиноререпторов (курареподобными пре-
паратами, бунгаротоксином) нарушает открытие Nа+ каналов на постсинапти-
ческой мембране. Фосфоорганические соединения (множество инсектицидов)
19
нарушают эффективность АХЭ и вызывают длительную деполяризацию пост-
синаптической мембраны. В клинике используют специфические блокаторы нервно-мышечного проведения: блокада холинорецепторов курареподобными препаратами, сукцинилхолином и другими конкурентными ингибиторами, вы-
тесняющими АХ с холинорецептора.
Миастения (myasthenia gravis) встречается примерно у 1 из 20000 людей.
При этом заболевании из-за неспособности нервно-мышечных соединений пе-
редавать достаточное количество сигналов с нервных волокон на мышечные возникает мышечный паралич. Это связано с дефицитом холинорецепторов на постсинаптической мембране (из-за их аутолитического разрушения) и поэтому возникает прогрессирующая мышечная слабость, вплоть до полной остановки мышечных сокращений (остановка дыхания). В этом случае используют блока-
торы АХЭ (неостигмин, физостигмин), что приводит к увеличению длительно-
сти связывания медиатора с меньшим количеством холинорецепторов и не-
сколько увеличивает амплитуду деполяризации постсинаптической мембраны.
Многие химические соединения (метахолин, никотин, карбахол) оказыва-
ют такое же влияние на мышечное волокно, как и ацетилхолин. Различие меж-
ду действием этих веществ и ацетилхолином заключается в том, что эти веще-
ства не расщепляются ацетилхолинэстеразой или разрушаются так медленно,
что их действие часто продолжается от нескольких минут до нескольких часов.
Это может приводить к длительной деполяризации, вызывая состояние мышеч-
ного спазма.
Контрольные вопросы
1.Из каких частей состоит нервно-мышечный синапс?
2.Какой медиатор используется для передачи ПД в мионевральном синапсе?
3.Какие рецепторы находятся на постсинаптической мембране нервно-
мышечного синапса?
4.Какую функцию выполняет фермент ацетилхолинэстераза в синапсе?
5.Из каких этапов состоит нервно-мышечная передача возбуждения?
20
6.Каким образом можно воздействовать на синапс фармакологическими пре-
паратами?
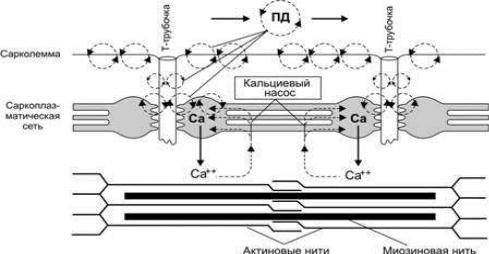
1.5.Механизм мышечного сокращения
Впроцессе сокращения мышечного волокна в нем происходят следующие преобразования:
А. Электрохимическое преобразование (рис. 9) - это последовательность
процессов, в результате которых потенциал действия плазматической мембра-
ны мышечного волокна приводит к запуску цикла поперечных мостиков.
Рис. 9. Распространение потенциала действия по сарколемме мышечного волокна и выброс ионов кальция из цистерн саркоплазматического ретикулума
Выделяют следующие этапы:
1.Генерация ПД.
2.Распространение ПД по Т-системе.
3.Электрическая стимуляция зоны контакта Т-системы и саркоплазмати-
ческого ретикулума, активация ферментов, образование инозитолтрифосфата,
повышение внутриклеточной концентрации ионов Са2+.
Б. Хемомеханическое преобразование:
В 1954 г. H.T. Huxley и A.F. Huxley независимо друг от друга для объясне-
ния мышечного сокращения сформулировали теорию скольжения нитей. Авто-
ры данной теории утверждают, что укорочение саркомера, а следовательно, и