Как проводят оценку?
Для этой цели служит определение содержания витамина в сыворотке крови, либо определение суточной экскреции метилмалоновой кислоты, которая возрастает при низкой обеспеченности организма кобаламином в десятки и сотни раз. Иногда применяется также метод нагрузки с помощью парентерального введения меченного по кобальту витамина В12.
Суточная потребность витамина b12, пищевые источники кобаламина
Синтез кобаламинов в природе осуществляется исключительно микроорганизмами. Животные и растительные клетки такой способностью не обладают. Основные пищевые источники витамина — печень, мясо (в нем кобаламина в 20 раз меньше, чем в печени), морские продукты (крабы, лососевые, сардины), молоко, яйца. У строгих вегетарианцев, исключающих из пищи не только мясные, но и молочные продукты, рано или поздно развивается В12-дефицитная анемия.
Суточная потребность — 3 мкг.
Продукты,богатые витамином b12






Химическое строение и свойства витамина h
Биотин был выделен в 1935 г. из яичного желтка. Свое название витамин получил от греч. bios — жизнь из-за его способности стимулировать рост дрожжей и бактерий.
Молекула витамина Н состоит из имидазольного и тетрагидротио-фенового колец, боковая цепь представлена валериановой кислотой.
N1-имидазольного кольца является местом карбоксилирования. Связываясь с ионом гидрокарбоната (НС03-), биотин становится кофер-ментом, называемым карбоксибиотином.

Биотин плохо растворяется в воде, но хорошо в спирте. Он устойчив при нагревании и в растворах слабых шелочей и оснований.
Биотин способен образовывать с авидином — гликопротеином белка куриного яйца — прочный комплекс, который не может расщепляться пищеварительными ферментами. Поэтому при частом употреблении сырых яиц прекращается всасывание присутствующего в пище биотина. Способность молекул авидина и биотина специфически связываться друг с другом используется в некоторых методах очистки в биотехнологии.
Метаболизм биотина (витамина h)
С растительной пищей витамин Н поступает преимущественно в свободном состоянии. Биотин животной пищи освобождается гидролазами от связи с различными белками и в свободном виде всасывается в тонком кишечнике. В кровяном русле биотин переносится альбумином и аккумулируется главным образом в печени. В тканях биотин находится в виде карбоксибиотинил-ферментов: СОО- группа валериановой кислоты карбоксибиотина ковалентно присоединена карбамидной связью к e-NH2-rpynne лизина, входящего в состав активного центра биотинзависимого фермента.
Выводится биотин в свободном виде с мочой и экскрементами, причем с последними его выводится больше, чем поступает с пищей. Объясняется это способностью микрофлоры кишечника синтезировать биотин.
Биохимические функции биотина
Витамин Н способствует усвоению тканями ионов бикарбоната (но не СO2) и активирует реакции карбокси-лирования и транскарбоксилирования в составе следующих карбо-ксибиотинил-ферментов:
Пируваткарбоксилазы — фермента, катализирующего АТФ-зависимое образование оксалацетата из пирувата и НСО3- .

Пируваткарбоксилаза является тетрамерным белком, несущим четыре молекулы биотина, каждая из которых связана с остатком лизина алофермента. Пируваткарбоксилазная реакция является наиболее важной анаплеротической реакцией, особенно в печени и почках (к анаплеротическим относятся возмещающие, пополняющие, реакции). Так, пируваткарбоксилаза восполняет запас оксалацетата, необходимый для функционирования цикла Кребса.
Пируваткарбоксилаза является важным митохонлриальным ферментом глюконеогенеза (новообразования глюкозы).
Ацетил-КоА-карбоксилазы — первого фермента в реакциях биосинтеза жирных кислот. Активная форма энзима представляет собой множество длинных мономерных нитей. При ферментативном катализе отдается карбоксильная группа бикарбоната ацетил-коэнзиму А с образованием малонил-КоА:

Пропионил-КоА-карбоксилат — фермента, участвующего в окислении жирных кислот с нечетным числом атомов углерода. При этом происходит стереоспецифический перенос активированной карбоксильной группы от карбоксибиотина к пропионил-КоА с образованием метил малонил-КоА:

Следует отметить, что ион бикарбоната может утилизироваться клеткой без участия биотина, как, например, это имеет место в карбомоилфосфатсинтетазной реакции при синтезе пиримидинов:
![]()
Бета-метилкротоноил-КоА-карбоксилазы — фермента, участвующего в реакциях окислительного распада лейцина.
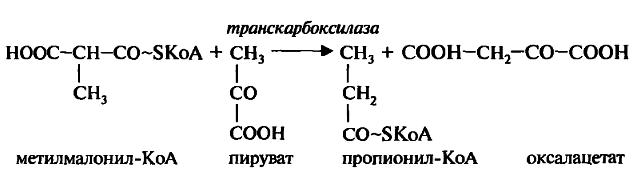
Метилмалонил-ЩУК-транскарбоксилазы — фермента, катализирующего реакцию транскарбоксилирования, а именно, обратимое превращение пирувата и оксалацетата (другие транскарбоксилазные реакции также протекают с участием биотина):