

3 курс / Патологическая физиология / Основы общей патологии
.pdf! |
|
331 |
|
клеток — вызваннойсамымиразнымипр чина |
|
минапример( ,в |
|
апоптозелимфоцитовподдействиемкакглюкокортикоидов,так |
|
|
|
вирусаСПИДа)Пути. патогенногодействияАКРпредставленына |
|
|
|
рисунке 37. |
|
|
|
Легкозамет,чтопрвсехивышеназванныхтьпроцесса |
|
х,когда |
|
гибельклетидетсактивнымучастиемсвободныхрадикалов |
|
|
|
отсутствуетпервичксиягипилинявляетсяа глубокой. |
|
|
|
ПоказаночтовысконцентрациикиеАКРмогутиндуцировать |
|
|
|
некробиоз,носредниемалыеконцтжеАКРхнтрации |
|
|
|
(наперек, ример |
исиводорода)способнывключатьпрограмму |
|
|
апопт.Прооксидантза2,3 |
-диметокси-1,4-нафтодхинонвмалых |
||
концентстимулируклетокп ацол ,вяхфсреднихрацию |
— вызываетнекроз. |
— |
|
запускаетапоптоз,ввысоких |
|
|
|
По-видимому,свободнорадикальнаягибельклетк |
|
и,доизвестной |
|
степени,альтернативнаописаннвышегипоксическому |
|
|
|
некр,хотяимеетбиозусниммногообщего,включаяпочти |
|
|
|
идентичныефинальныестадии. |
|
|
|
Какивслучаекальцием,граньмеждустимуляцией |
|
дьбаклеткипри |
|
повреждением — временнаяиколичественная,су |
|||
активацииАКР |
— этов балпрос.Таобнсаким,равновесиезом |
|
|
междугенериинациейАКРктивациклеткеможетй |
— иэтофундаментальная |
||
устанразнвлиуровматьсяне |
|
||
неспецифичсоставляющаяаяклеточнойр , тивности |
|
|
|
модуклирующаяет |
очныеотвнаразнообразныетыконкретные |
|
|
стимулы.Биор,пегуляторыречисленныевышетаблице4,всез исключениятакилиинзатрбапрооксидантныхгиваютланс антиоксисткл.идантныхетким
АНТИОКСИДАНТНЫЕМЕХАНИЗМЫ КЛЕТОК
Антиоксиданты — этом леку |
лы,обладающиелабильным |
водородныматомомснеспареннымэлектроном: |
|
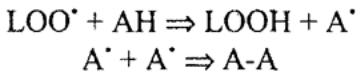
! |
332 |
где АН — антиоксидант, А-А его стабильный несвободнорадикальный продукт.
Множество антиоксидантов, вырабатываемых клетками и поглощаемых извне в качестве полностью, либо частично незаменимых соединений сдерживают клеточное «атомное оружие», препятствуя длительному существованию высоких концентраций АКР. Кроме того, избыток АКР может секвестрироваться в пероксисомах. Антиоксиданты — не просто набор веществ. Они способны восстанавливать друг друга и представляют собой антиоксидантные системы клеток.
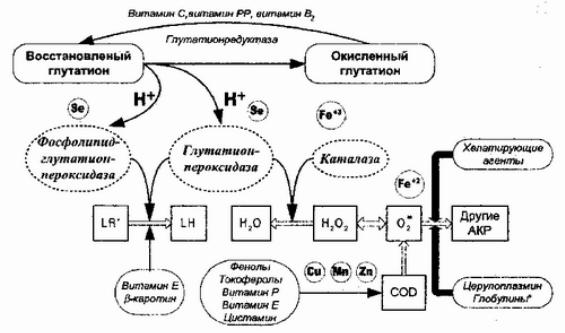
Основными разрушителями АКР служат ферменты каталаза, супероксиддисмутаза, глютатионпероксидаза, фосфолипид- глютатионпероксидаза и глютатионредуктаза.
По П. Хорнсби и Дж. Кривелло, имеются три класса антиоксидантов (1983):
Каталаза и глютатионпероксидаза — это энзимы предупредительного действия поскольку они восстанавливают АКР (перекись водорода), провоцирующую цепной свободно- радикальный процесс, до неактивного состояния, Супероксиддисмутаза — фермент-прерыватель цепной реакции. Она превращает при [190] наличии восстановительных эквивалентов, супероксидный анион, способный, как показано выше, формировать наиболее активные АКР, в менее активную перекись водорода, разрушаемую каталазой. Субстратами — прерывателями цепной реакции служат фенолы (например, токоферол) и амины (например, цистамин).
Третья разновидность антиоксидантов — хелатирующие агенты, способные связывать железо и другие металлы- катализаторы и разветвители цепных свободнорадикальных реакций (например, десферол и унитиол).
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
333 |
Всеупомянутыеэнзиихзоэнзимыявляются металл.Всоихставактиферментамицентровходятных микроэлементы.
Глутатифосфолипиднпероксидаза |
|
-глутатионпероксидаза, |
|||
утилизующиеэтотметаболитдляинактивации |
|
|
|
перекисиводорода |
это |
липоперекисейобразованиемспиг товойуппы |
|
|
|
— |
|
селеносодферм.Различныетканевыержащиентыизоферменты |
|
|
|
|
|
суперокссодержатц,инкмарганецддлиимедьсмутазы. |
|
|
|
|
|
Митохизоиспользуетаяндриформамарга,цитозольныеец |
|
|
|
|
|
— цинк медь. |
|
|
|
|
|
Катаявляетсяпероксисомальнымазажелезозависимым |
|
|
|
||
металлоферментом. |
|
|
|
|
|
Глантиоксиданвныесубстратыклетокные |
|
|
— это тиоловые |
||
соединения. |
|
|
|
|
|
Книмотносятся |
глютатион, цистеин, Д-пеницилламин. |
|
|||
Глютатион — важнейшкомпонентантисистемокйидантных |
|
|
|||
печени,сердца,моз,легкихклеткровиа.Показанаего |
— пригемолизе,длянейронов |
— |
|||
защитнаяродэритроцитовль |
|
||||
приинсультенейродегензабол, альвеоцитоввныхяванрях |
|
|
— |
при |
|
— приотравозоном,длякарденииомиоцитов |
|
|
|||
миокардиодистрофиях.Глютат |
иоблнрадиопротекторнымиает |
|
|||
свойствами. |
|
|
|
|
|
Дляобразовнеобходяглютатамиоцистеинокислота,ма |
|
|
|
|
|
пработеглутатионовыхпероксидонпрев азщается |
|
|
|
|
|
дисульфид. |
|
|
|
|
|
Другаягруппавеществ,испокльзуеетканашегомыхи |
|
огостресса |
— этовитамины. |
|
|
организмадлязащитыотокислительн |
|
|
|
||
Реактивируетглутатиондисульфидфермент, ионре уктаза |
|
|
|
||
зависотНАДФНиаскорбиновоймыйкислоты.Сле,довательноля |
|
|
[191] дисульфида |
||
восстановленияреактивглютатиз ц иона |
|
|
|
||
нужнынетольмик,рониовитаэлеР меиныты |
— |
такжеЕиВ |
|
РиС,а,по |
|
некоторымсведениям |
2 Однако,аскорбиновая |
|
|||
кислотаамбивалентнаокислительно |
|
|
-восстановительныхреакциях |
|
|
! |
334 |
и способна оказать и прооксидантные эффекты, в частности, ускоряя восстановление железа, усиливающего свободнорадикальные процессы. По Хорнсби и Кривелло, прооксидантные эффекты витамина С преобладают при его малых дозах, а антиоксидантные — при больших, что, в какой-то степени, оправдывает рекомендации Л. Полинга замедлять старение и предупреждать повреждение клеток, в частности, при вирусных инфекциях и атеросклерозе, мегадозами аскорбиновой кислоты (1983). Интересно, что, по мнению Б. Н. Эймса и соавторов (1981), у приматов таким же, как витамин С, амбивалентным действием на редокс-состояние клеток обладает мочевая кислота (см. также выше «Диатезы»).
Особенно тесная взаимозависимость существует между селеном и витамином Е, которые оба служат для инактивации липоперекисей. Витамин Е является сильнейшим антиоксидантом, так как ловит свободный электрон и не участвует в дальнейшей цепи. Протективное действие токоферола особенно выражено в отношении клеточных мембран. Классические эксперименты Дж. Блэнда с человеческими эритроцитами показали, что 10- дневный прием 600 ME α-токоферола ежесуточно делает плазматические мембраны 95% красных кровяных телец испытуемых резистентными к тому окислительному стрессу , который до курса лечения вызывал стопроцентный гемолиз.
Активность токоферолов восстанавливается витамином С, как и активность системы глютатиона.
Таким образом, в системе глутатиона взаимодействуют витамины, микроэлементы и серосодержащие аминокислоты. Упомянутые витамины и микроэлементы, а также полифенолы (биофлавоноиды), б-липоевая кислота и в-каротин действуют в комплексе и составляют антиокислительный резерв клеток, определяющий их резистентность к свободно-радикальному повреждению.
Многие пищевые продукты содержат значительные количества этих ингредиентов и способны насыщать ими организм.
Поэтому, питание, оптимально обеспечивающее потребности
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
335 |
клетоквантиоксидантах,является внастоящеевремяпредметом наиболеентенсивныхразработокдиетологии.
Аутоокислипопротяводяетсямеханизмовидов атерогенеза,свободноповреждениеДНКспособноадикальное вызыватьканцерогенез,поэтомуантиоксидантыпроявляют протективноедействиеотношенииат ропухолевыхсклероза заболеваний.
Дефицитнезамсо ,отднинмыхкэтоящихсяенс ,йстеме связанразвитиеммногихзаболеваний,сопровождаемых клеточнымокислительнымповреждением.
Так,область,эндемичнаяпогеохимическ |
|
|
омудефицитуселенав |
|
провиЖэКНР(н)отличаетсяцииьсуповышеннчастос тбой |
болезньКишэн |
),аВосточнаяФинляндия |
|
|
миокардиодистрофии( |
|
|||
имеетсамоенизкоевмирегеохимическоесодержаниеселена |
|
|
|
|
рекордномалуюпродолжительностьпредстоящейжизни |
|
|
|
|
сорокалетнихмужчин. |
|
|
|
|
АмериканавтУоллбнсавязькиймеруждуил |
|
|
|
|
муковидефицитомсцидозомелуб наре енных |
|
|
|
|
новорожденных. |
|
|
|
|
Тиоловымиантиоксидслужатбогатыентами |
|
|
|
|
сульфгидрильнымигруппамицитозольныебелкитиоредоксин( ),а |
имакрофагпроисхождения,такиельного |
|
||
такжесывороточныебелк |
|
2- |
||
какцерулоплазмин,С |
|
-реактивныйбелок,гаптоглобин,α |
||
микроглобул,амилоидА.Перечбелкисисленныетезируются |
|
-1и |
||
макразличнойофаглоквответминанзацииерлейкин |
|
|
||
ряддругихмедиатороввосп.Т кимления |
|
|
бразом,осуществляется |
|
попредоыткаорганизмхранозяотнегативныхнапоследствийть |
|
|
|
|
собственокислительногоудара, анпоагентамсимого, |
|
|
|
|
вызвавшимвоспаление. |
|
|
|
|
Именновследусилениясинттвиесыворзаглобулиновточных |
рошоизвестнмедикукаждому |
|
||
— антиоксидантовпроисходит |
|
|
(СОЭ). |
|
[192] увеличениескорости |
|
реакцииоседанияэритроцитов |
||
Всвязиун вехарактерос льнымссатриваемыхеханизмов, СОЭускоряетсяприширокомкругеинфекций,воспалений
! |
336 |
иммунопатологических процессов разной этиологии.
Белки, усиленно синтезируемые при воспалении объединяют под названием «положительные глобулины острой фазы» (см. ниже разделы «патофизиология Воспаления» и «Преиммунный ответ»). Типичный представитель этой группы протеинов — церулоплазмин является феррооксидазой и окисляет двухвалентное железо до трехвалентного без образования свободных радикалов.
Особую роль в антиоксидантной защите играет трансферрин — отрицательный глобулин острой фазы, содержание которого в крови при воспалениях и инфекциях снижается.
Он захватывает трехвалентное железо и может переносить его в клетки.
Как уже отмечалось выше, адаптивная ценность гипоферремии при остром ответе на инфекцию экспериментально доказана и связана с каталитической ролью двухвалентного железа в системе генерации АКР.
Эта форма железа чрезвычайно цитотоксична, поскольку участвует в реакции Фентона и в реакциях разветвления цепей окисления мембранных липидов. При лихорадке изменения в синтезе глобулинов острой фазы ограничивают аутоокислительный эффект железа.
Кора надпочечников обладает очень высокой активностью цитохром-Р-450-зависимых оксидаз, и поэтому продукция АКР в этих органах крайне активна, особенно, в условиях стресса. Именно поэтому надпочечник обладает исключительно высокими потенциями антиоксидантных механизмов (рекордные количества аскорбиновой кислоты и витамина Е, очень высокий уровень супероксиддисмутазы). Хорнсби и Кривелло считают редокс- состояние адренокортикоцитов важным фактором, влияющим на интенсивность стероидогенеза и на морфогенез коры надпочечников, в частности, на апоптотическую гибель и эскалаторное перемещение клеток из зоны в зону, а также на смену паттерна синтезируемых клетками различной локализации кортикоидов. Следовательно, АКР влияют не только на клеточные
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
|
337 |
|
|
процессыитканевойответнаповреждение |
— воспаление.Они |
|||
существенныидляразвисисответовияемныхнаповреждение |
|
|
— |
|
реакцостройфазыистреисм.ниже(соответствующиеса |
|
|
|
|
разделы). |
|
|
|
|
Итак,свободныерадикалывсегдаигропределеннуюютроль |
|
|
|
|
приклеточнгибе.Врядес особенно,иучаевй,прадиационной |
|
|
|
|
травме,отравленияххлорорганическимисоединениями |
|
|
|
|
воспаихвкладменииханизмыповрежденияклетокявляется |
|
|
|
|
определяющим. |
Окислительно-восстановительныйбалансклеток |
|||
зависитотэкспрегени освсииубстртавкинезаменимыхтов |
|
|
|
|
компондиетыпредставляетнтовсобойоднуизглавных |
|
рис. 38). |
||
неспецифичсоставляющихихклеточнойр (тивности |
|
|||
|
|
|
|
|
Рис. 38Антиоксид. механизмыорг. CODизматные |
— |
супероксиддисмутаза. |
|
МЕХАНИЗМЫАПОПТОЗА
Явления запрограммированнойклеточнойгибели |
известны |
|
ужеболет100В.( |
Флеминг, 1885),нооставалисьтени«» |
|
некробиотическихпро |
цессов,которыенапротяжениидесятилетий |
|
изучалисьнамногоболееактивно,чемпрограммируемаягибель.
! |
338 |
Этот вид клеточной гибели представляет собой важнейший интегральный компонент эмбриогенеза, морфогенеза и роста тканей, а также гормонозависимой инволюции. Он, наряду с лизосомальной аутофагией (см. выше) участвует в механизмах таких клеточных адаптации, как атрофия (уменьшение размеров клеток и числа функционирующих структур в них при сохранении жизнеспособности клетки) и гипоплазия (уменьшение органа вследствие уменьшения числа клеток в нем при сохранении его жизнеспособности).
Так например, одним из авторов этой книги было показано, что инволютивные изменения в коре надпочечников после гипофизэктомии тормозятся актиномицином Д, а значит, представляют собой активный процесс реализации [193] некой программы саморазборки клеток (А. Ш. Зайчик, 1978).
Для обозначения процесса запрограммированной клеточной гибели, морфологически и патохимически отличного от некробиоза, Н. Уолкер в 1968 году предложил термин «апоптоз». Основатели учения об апоптозе, в частности, Дж. Керр и соавт., считали понятия «запрограммированная клеточная гибель» и «апоптоз» равнозначными. В последнее время имеется тенденция применять первый термин к процессам устранения клеток в раннем онтогенезе, а понятие апоптоз относить только к программируемой гибели зрелых дифференцированных клеток. Так, указывают на наличие аутофагии и отсутствие разрывов ДНК при эмбриональной клеточной гибели, в отличие от апоптоза зрелых клеток.
Вопрос о соотношении некробиоза и апоптоза и о приуроченности этих механизмов к естественной либо насильственной гибели клеток нуждается в обсуждении.
Было бы упрощением сказать, что апоптоз это, исключительно, процесс естественной гибели клеток, а некробиоз — насильственной. Деление на эти два процесса далеко не абсолютно. Выше, обсуждая паттерны некробиоза, мы уже много раз вынуждены были упоминать об апоптозе, так как между этими процессами много общего. Дело в том, что в ответ на минимальное повреждение или повреждение, не вызывающее быстрого развития
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
339 |
|
|
|
|
глубокойгипоксиивыраженногоэнергодефицита,клеткимогут |
|
|
|||
включатьспециальнуюпрограммусамоуничтоженияреагировать |
|
|
|||
апоп.Вэтослучае,назомпри, действиимериониз рующего |
насильственна,но |
||||
излученияиливирусаСПИДа,смертьклетки |
|
|
|
||
механизмеенекробиотический,апоптотический. |
|
Тельца |
|||
Каунсильмена обнаружиприв русногепвпечениатитеые, м |
|
|
|||
представляютсобойрезультапоптозагепатподоцитов |
|
— также |
|||
воздействиемвирус |
-индуцированногоповреждения.Это |
|
|||
насильственнаягибель,номеханиеенесвязанм |
|
|
|||
быстропргипипозволяксигресклуспткейирующейеть |
|
|
|||
включитьпрограммусаморазборки.Неподлежитсомнению |
-мишенейподвоздействием |
|
|||
насильствхараклетокргибелинный |
|
|
|
|
|
факторанекрозаопухолей.Однако,не |
|
|
|
смотрянасвоекатегоричное |
|
назва,данныйбиорегуляториевызыватаргеклетканеных |
|
|
|||
тольконекроз,ноиапоптоз.Приреализациинекробапоптоза |
|
|
|||
функционмногиеобщмеханизмы,напримерруют,увеличение |
анногокальция |
|
|||
цитоплазматическойконцеионтрацииизиров |
|
||||
образсвободныхактваниекислородныхвныхрадикалов.Более |
|
[195] и |
|||
того,прибольшейсиле |
[194] Рис. 39. Механизмыапоптоза. |
||||
|
|
|
|
|
|
интенсивностидействияапоп можетимулогенныйвызвать некр,очевиднобиоз,вследствиетого,чтопрогрессирующий энергодефицитдаетвозможностиклеткамреализовать энергетдорогую« » инамикуческиапоптоза.
Авторампредстапринципиальноважнымляется,чтоапоптоз,в |
остьклеточной |
противопнекр,эторазнложностьбизувидн |
|
гибе,еслиможнотаквыразиться,безсопутствующегоскандала. |
|
Еслинекробивсегдасопровождаетсязсвобождением |
— ивсистемный |
окружающуюткань,примассивномпоражении |
|
кровоток,медиатороввосп,ч лест,линпостиияидныхродуктов |
|
деструкцииклеточныхмембран, апоптозпротекаетбе |
|
лейкоцдемаркацииперифоктарнвоспй,таккакалеьнияого |
|
егомеханипозволяетизбежатьзначительногом выделения |
|
медиатклеточногоп роввреждения.Изданиегоду1996 |
освященнойапоптозу |
всеобъемлющеймонографии,п |
|
(«Программированнаяклеточнаягибель»подред.профессора |
|
В. С. Новикова)облегчнашузаделаетвозможнымчу |
|
охарактеризоватьданкнлойнаигешьболеещие |
|
! |
340 |
патофизиологически важные аспекты этой проблемы.
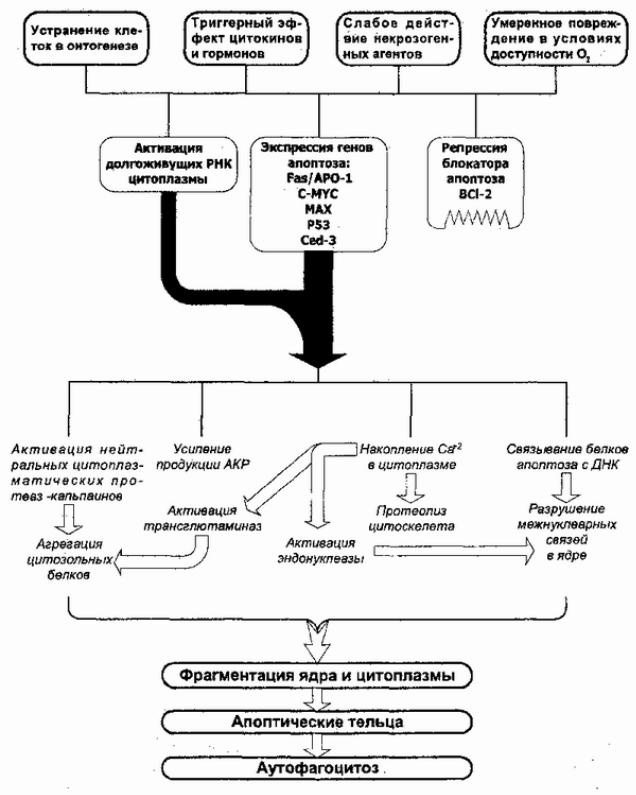
Рис. 39. Механизмы апоптоза. Пояснения в тексте.
В. С. Новиков и соавторы выделяют следующие основные
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/