

2 курс / Биохимия / Книжки и сборники / Biokhimia_dlya_chaynikov
.pdfгликогена прерывается. Угнетение синтеза гликогена также означает, что ор- ганизму доступно больше глюкозы для совершения немедленного действия, например бегства от разъяренного питбуля.
Прикрепление гормона к рецепторному участку провоцирует ответ, который
будет повторяться до тех пор, пока не будет разорвана связь с гормоном. На-
пример, если ответом будетобразование молекулы, то запускающий этот про-
цесс гормон будет вызывать образование не одной молекулы, а непрерывного потока молекул. Такой непрерывный синтез не прекращается до тех пор, пока
гормональная молекула будет оставаться на рецепторе. Согласно этому меха- низму одна молекула гормона можетиндуцировать образование тысяч других молекул. Этот процесс, называемый амплификацией, приводит к более эффективному результату, чем можно судить по низкой концентрации гормона.
Стероидное регулирование
В отличие от механизма вторичных мессенджеров в стероидном регулиро-
вании гормоны проходят через мембрану и заходят в клетку. Этот механизм
применим к некоторым другим гормонам, например вырабатываемым щито-
видной железой.
Впервые этой моделью было описано действие эстрадиола на ткани матки
у млекопитающих. Эстрадиол или эстроген проходит через клеточную бис-
лойную мембрану, где связывается с эстроген-рецепторным белком. При этом ковалентная связь не создается, а вместо этого индуцируется конформацион- ное изменение белка. Изменение формы позволяет белку пройти через обра- зованный “ проем” внутрь клеточного ядра. Затем в клеточное ядро проходит гормон-протеиновый комплекс, где связывается с определенным участком на
хромосоме. Такое объединение с хромосомой стимулирует транскрипцию для производства иРНК, которая выходит из ядра и обеспечивает синтез белковых молекул с помощью механизма трансляции.
У стероидной системы есть три отличительных преимущества перед меха- низмом вторичных мессенджеров. Во-первых, при стероидном регулировании
гормон проходит непосредственно через клеточную мембрану. Во-вторых, в цитоплазме (внутриклеточной жидкости) присутствует готовый стероидный молекулярный рецептор. И наконец, гормон действует на хромосомном уровне.
210 ЧАСТЬ 3 Углеводы, жиры, нуклеиновые кислоты и другие...
г
-
ШХ |
X XX |
: |
|
' |
|
|
шш |
|
I
г |
: : |
iSSt |
- |
|
|
1 |
|
|
рге-
к:
и
метаболизм
Для совершения действия необходима энергия. В этой части мы сфокусируем свое внимание на том, как клетки получают и расходуют энергию. Рассмотрим энергетические потреб-
ности организма и попытается понять, куда и с какой целью тратится энергия. Основным действующим лицом этой гла-
вы будет наш лучший друг и незаменимый помощник —
АТФ. Познакомимся с циклом лимонной кислоты (циклом Кребса), а в конце — с химическими реакциями, связанными с образованием азотных соединений.


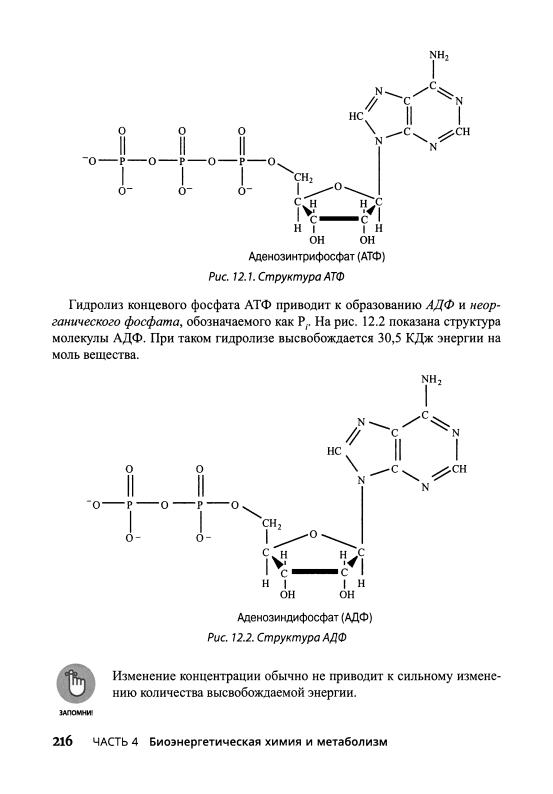
Гидролиз концевого фосфата АДФ вызывает образование АМФ и неор- ганического фосфата, обозначенного как Р.. Структура АМФ приведена на рис. 12.3. В процессе такой реакции высвобождается 30,5 КДж энергии на моль вещества. (С биохимической точки зрения эта реакция имеет меньшую
значимость, чем гидролиз АТФ с получением АДФ.)
|
|
|
|
|
|
|
|
— |
|
NH2 |
|
|
|
|
|
|
|
N |
|
|
|
|
|
|
|
|
|
|
// |
^С |
N |
|
|
|
|
|
|
|
|
/ |
|||
-О |
Р |
О |
|
|
НС |
\N |
|
N ^ |
||
|
|
|
|
|||||||
|
О |
|
|
|
|
|
|
|
|
СН |
|
О - |
СН2 |
о |
ц |
|
|
|
|||
|
|
|
|
|
|
|||||
|
|
1 |
|
|
|
|
|
|||
|
|
С |
|
с |
I |
|
|
|
||
|
|
н |
| |
|
|
I |
Н |
|
|
|
|
|
|
он |
|
он |
|
|
|
||
Аденозинмонофосфат (АМФ)
Рис. 12.3.Структура АМФ
Непосредственное превращение АТФ в АМФ также возможно — в резуль-
тате отщепления пирофосфата (Р^у") от фосфатной цепи. В биохимии пиро-
фосфат обозначается как РР.. В процессе такой реакции образуется энергии
немного больше, чем при простом гидролизе с освобождением Р;. (около 33,5 КДж на моль вещества). В физиологических условиях фосфатная часть АТФ и АДФ образует комплекс с ионами магния. При определенных обстоя-
тельствах ионы магния Mg2+ могут быть замещены двухвалентными ионами марганца Мп2+. На рис. 12.4 показано, как магниевые комплексы включается в
структуры АТФ и АДФ.
Выделение последнего фосфата влечет за собой потерю минимального количества энергии: 14,2 КДж на моль"1. При таком гидролизе расщепляется эфирная, а не ангидридная связь. В общем случае гидролиз эфирной связи дает
меньше энергии, чем гидролиз ангидридной связи.
ГЛАВА 12 Жизнь и энергия 217
|
|
|
|
|
|
|
|
|
|
|
NH2 |
|
|
|
|
|
|
|
|
N |
С /ЧN |
||
|
|
|
|
|
|
|
|
// |
|
|
|
о |
|
о |
|
о |
|
|
НС\ |
^с |
N^СН |
||
'0 о |
о |
р |
о |
о |
сн2 |
о |
|
N |
|||
0' |
Mg |
о |
|
о |
нН I |
У |
|
|
|
||
|
\ |
|
|
|
|
|
\ |
|
|
|
|
|
|
2+ |
|
|
|
|
|
|
|
||
|
|
|
|
|
I |
Н |
|
|
|
||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
он |
он |
|
|
|
||
|
|
|
|
Аденозинтрифосфат (АТФ)-Мд2+ |
|
|
|
||||
|
|
|
|
|
|
|
|
|
NH2 |
|
|
|
|
|
|
|
|
N |
|
|
^ |
|
|
|
|
|
|
|
|
// |
С |
|
N |
||
|
|
|
|
|
|
|
|
|
|
||
|
с |
|
О |
|
|
НС\ |
N |
|
N |
^ |
СН |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||
О |
|
о |
э |
о |
|
|
|
|
|
|
|
D |
|
|
|
|
|
|
|
||||
|
|
|
|
сн2 |
о |
|
|
|
|
|
|
|
о |
|
о |
нН I |
Vr |
|
|
|
|
||
|
|
|
|
|
|
|
|||||
|
|
Mg 2+ |
|
|
|
|
|
|
|||
|
|
|
|
|
? |
Н |
|
|
|
|
|
|
|
|
|
он |
он |
|
|
|
|
|
|
Аденозиндифосфат (АДФ)-Мд2+
Рис 12.4. Магниевые комплексы в АТФ и АДФ
Все познается в сравнении:
молекулы, связанные с АТФ
Мало какие биомолекулы в состоянии предоставить столько энергии,
сколько высвобождается при гидролизе АТФ. Одна из них ГТФ. Сравнимое
количество энергии предоставляют некоторые другие молекулы. В табл. 12.2 приводится сравнение некоторых высокоэнергетических молекул с АТФ, а на рис. 12.5 показаны их структуры.
218 ЧАСТЬ 4 Биоэнергетическая химия и метаболизм
Таблица 12.2. Энергия (AG0 / b выделяемая некоторыми
высокоэнергетическими молекулами по сравнению с АТФ
АТФ |
30 |
5 |
|
, |
|
Фосфоаргинин 32,2
Ацетилфосфат 43,3
Фосфокреатинин 43,3
1,3-бифосфоглицерат 49,6
Фосфоэнолпируват 62,2
Фосфоэнолпируват, 1,3-бифосфоглицерат и ацетилфосфат задействованы в
переносе и сохранении химической энергии. Фосфоаргинин и фосфокреати-
нин — это важные молекулы, предназначенные для хранения энергии, полу- ченной в процессе обмена веществ. Фосфокреатин накапливается в мышцах и может быстро превращаться в АТФ, чтобы предоставить энергию мышцам для сокращения. Выработка фосфокреатинина происходит при высокой концентра- ции АТФ, когда необходимо превысить дефицит энергии в 12,8 КДж на моль вещества. Обратный процесс переноса фосфата от АТФ к АДФ возникает при низкой концентрации АТФ. Фосфоаргинин ведет себя аналогично у некоторых
беспозвоночных (у них “ стальные” мышцы!).
Семейство нуклеозидтрифосфатов
Как мы уже говорили, основная молекула-переносчик энергии — это АТФ.
Но другие нуклеозидтрифосфаты, такие как ЦТФ, ГМФ, ТТФ и УТФ также могут выполнять функцию переноса энергии. Кроме того эти пять молекул
частично обеспечивают необходимой энергией процесс синтеза ДНК и РНК. Все нуклеозидтрифосфаты дают примерно одинаковое количество энергии.
(Заметьте, АТФ необходима для синтеза других нуклеозидтрифосфатов.)
Биосинтез рибонуклеозидтрифосфата с общим названием НТФ начина- ется с выработки соответствующего монофосфата — НМФ. Для поэтапного
добавления следующих двух фосфатных групп требуется два узкоспецифич- ных фермента. Такими ферментами являются нуклеозидмонофосфаткиназа и нуклеозиддифосфаткиназа. (Термином киназа обозначаются трансферазные ферменты, которые переносят фосфатную группу нуклеозидтрифосфата.) На рис. 12.6 приведен общий вид такой реакции.
ГЛАВА 12 Жизнь и энергия 219
H NH2
|
I - |
|
- |
|
- |
- |
- |
|||
|
HN |
NH |
сн2-сн2 сн2 |
сн2 |
сн |
|||||
|
О |
|
|
|
|
|
|
|
|
с=о |
|
|
|
|
|
|
|
|
|
|
|
о |
P=0 |
|
|
|
|
|
|
|
|
он |
|
—О" |
|
Фосфоаргинин |
|
|
|
|
|||
|
|
|
о |
|
о |
|
|
|
|
|
|
сн3-с—о—О"—о |
|
|
|
||||||
|
|
|
Ацетилфосфат |
|
|
|
|
|||
|
|
|
NH |
|
|
О |
|
|
|
|
|
HN- |
|
|
—N |
СН2-С О |
|
||||
|
о- |
|
о |
СН3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
О-Фосфокреатинин |
|
|
|
||||||
|
|
|
|
|
о |
|
|
|
|
|
|
|
н2с |
сн |
с |
|
|
|
|
||
|
|
|
о—он |
—о |
|
|
|
|
||
|
О |
|
|
|
^О |
|
|
|
||
|
|
"— |
|
|
— |
” |
|
|
|
|
|
0=р |
0“ “О |
|
Р=0 |
|
|
||||
|
1,3-бифосфоглицерат |
|
|
|
||||||
|
|
|
|
|
/о |
|
|
|
|
|
|
|
|
СН2=С—с\/ о |
|
|
|
|
|||
|
|
|
|
о |
|
|
|
|
|
|
|
|
|
0: |
р - |
О- |
|
|
|
|
|
|
|
|
|
О" |
|
|
|
|
|
|
Фосфоэнолпируват
Рис. 12.5.Структуры некоторых
высокоэнергетических молекул
220 ЧАСТЬ4 Биоэнергетическая химия и метаболизм