

Ответы на зачёт по НС
.pdfВестибулярные рефлексы неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их. Вестибулярные рефлексы не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой.
Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические. Статические рефлексы связаны главным образом с возбуждением рецепторов преддверия перепончатого лабиринта и обеспечивают поддержание позы и равновесия тела при самых разнообразных его статических положениях в пространстве. Так, например, если кошку с фиксированной относительно туловища головой поворачивать в пространстве, то в зависимости от положения головы будет наблюдаться различное распределение позного тонуса. В том случае, когда угол между плоскостью горизонта и плоскостью ротовой щели животного составляет 45°, разгибательные мышцы напрягаются максимально. Если же указанный угол равен 135°, наблюдается наименьшее значение тонуса разгибателей.. Между крайними положениями имеется ряд постепенных переходов в степени развития экстензорного тонуса. Это пример тонического вестибулярного рефлекса положения, который осуществляется при участии нейронов латерального вестибулярного ядра (Дейтерса) и идущего от него в спинной мозг преддверно—спинномозгового пути.
К числу статических вестибулярных рефлексов относят рефлексы выпрямления, направленные на переход животного из неестественной позы в обычное для него положение. В качестве наглядного примера можно привести переворачивание кошки в воздухе при падении спиной вниз. За очень короткое время падения животное успевает занять нормальное по отношению к гравитационному полю положение и упасть сразу на все четыре лапы. В процессе переворачивания выпрямительные рефлексы совершаются в определенной последовательности. Сначала за счет вестибулярного выпрямительного рефлекса восстанавливается нормальное положение головы — мордой вниз. Затем изменение положения головы возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный рефлекс, в результате которого вслед за головой туловище также возвращается в нормальное положение.
Таким образом, в естественных условиях вестибулярные рефлексы выпрямления дополняются шейными, причем ведущую роль в их осуществлении играет положение головы, где расположены дистантные рецепторы.
Следующая группа вестибулярных рефлексов — статокинетические — характеризуется тем, что она направлена на поддержание позы при изменении скорости движения животного. Эти рефлексы связаны с возбуждением рецепторов полукружных каналов, которое имеет место при наличии в каналах тока эндолимфы. Обычно направленность статокинетического рефлекса зависит от того, какой полукружный канал раздражается при ускорении. Например, при замедлении поступательного движения возбуждаются рецепторы сагиттального канала и, соответственно, рефлекторные изменения мышечного тонуса будут наклонять туловище вперед, как бы сохраняя равномерное движение.
Ускорение при вращении тела в горизонтальной плоскости возбуждает рецепторы горизонтального полукружного канала и вызывает рефлекторную реакцию глазодвигательного аппарата — горизонтальный нистагм. Суть этой реакции состоит в том, что в момент ускорения вращения глаза движутся в сторону, противоположную направлению вращения. Затем, достигнув крайнего отклонения, глаза быстро
перемещаются обратно в направлении вращения и таким образом в поле зрения попадает другой участок пространства. Нистагм способствует сохранению нормальной зрительной ориентации и обычно используется в диагностических целях для проверки нормального функционирования вестибулярного аппарата. Плоскость нистагма совпадает с плоскостью ускорения, в связи с чем кроме горизонтального нистагма можно встретить вертикальный, диагональный и круговой нистагмы .
К числу статокинетических вестибулярных рефлексов относятся также и так называемые лифтные рефлексы, которые проявляются в увеличении тонуса мышц разгибателей при линейном ускорении вверх и в повышении тонуса сгибателей при линейном ускорении вниз.
Произвольные движения или условные двигательные реакции возникают при индивидуальном развитии и накоплении новых навыков. Условные рефлексы формируются на основе безусловных с участием высших отделов головного мозга. Функционирование нервной системы проявляется в виде непрерывно возникающих ответных реакций на раздражающие факторы внешней и внутренней среды. Функциональной единицей нервной деятельности является рефлекс как ответная реакция нервной системы на раздражение.
Двигательный (корково-мышечный) путь
Общая характеристика.
Выполнение произвольных движений связано с выработкой плана и программы действия в двигательном анализаторе коры. Кроме того, необходимо, чтобы сформировавшийся здесь импульс произвольного движения достиг соответствующих мышц-исполнителей. Такое проведение нервного импульса от двигательного анализатора коры до поперечнополосатой мускулатуры осуществляет корково-мышечный или пирамидный путь. Данный путь двухнейронный – состоит из центрального и периферического двигательных нейронов.
Центральный двигательный нейрон образован телами нейронов преимущественно прецентральной извилины. Совокупность всех центральных двигательных нейронов
называют пирамидной системой. Тела этих клеток располагаются в V слое прецентральной извилины и парацентральной дольки. Впервые они описаны киевским анатомом В.А. Бецом в 1874 г., их называют гигантские клетки Беца. Существует четкое соматическое распределение этих клеток. Находящиеся в верхнем отделе прецентральной извилины и парацентральной дольке клетки иннервируют нижнюю конечность и туловище, расположенные в средней ее части – верхнюю конечность. В нижней части этой извилины находятся нейроны, посылающие импульсы к лицу, языку, глотке, гортани, жевательным мышцам. Таким образом, тела нейронов осуществляющие иннервацию отдельных мышечных групп, имеют расположение, обратное расположению частей человеческого тела. Другой особенностью двигательных областей коры является то, что площадь каждой их них зависит не от массы мышц, а от сложности и тонкости выполняемой функции. Особенно велика площадь двигательной области кисти и пальцев верхней конечности, в частности большого, а также губ, языка. Пирамидные клетки расположены не только в прецентральной извилине, также они имеются в задних отделах лобных извилин. Аксоны центральных двигательных нейронов направляются для образования синапса со вторым периферическим двигательным нейроном. Аксоны клеток, расположенных в верхней и средней третях прецентральной извилины носят название
кортико-спинальный (корково-спинномозговой) путь. Соответственно, аксоны клеток,
лежащих в нижней трети прецентральной извилины, образуют кортико-нуклеарный (корково-ядерный) путь. Аксоны всех этих нервных клеток направляются вниз и кнутри, приближаясь друг к другу, и составляют лучистый венец (corona radiata). Затем пирамидные проводники собираются в компактный пучок, образующий часть внутренней капсулы (capsula interna). Внутренняя капсула – это узкая пластинка белого вещества, расположенная между подкорковыми образованиями, имеющая вид тупого угла. Кортикоспинальный путь образует передние 2/3 задней ножки внутренней капсулы, волокна кортико-нуклеарного пути формируют колено внутренней капсулы. Далее волокна пирамидных клеток проходят в основании ножек мозга, варолиева моста. 9 В пределах мозгового ствола кортико-нуклеарные волокна совершают частичный надъядерный перекрест: часть волокон переходит на противоположную сторону и образует синапсы с нейронами двигательных ядер соответствующих черепно-мозговых нервов. Другая часть остается на своей стороне, образуя синаптические связи с клетками ядер этой же стороны. Исключение составляют ядро XII нерва и нижняя часть ядра VII нерва – на них замыкаются волокна корково-нуклеарного пути только противоположной стороны (т.е. данные волокна совершают полный перекрест).
Таким образом, мышцы языка и нижняя половина мимических мышц лица получают только одностороннюю корковую иннервацию от прецентральной извилины противоположного полушария, а другие мышцы (глазодвигательные, жевательные, верхняя группа мимических, мышцы глотки, мягкого неба, голосовых связок, гортани, шеи) обеспечены двусторонней корковой иннервацией. Поэтому одностороннее поражение кортиконуклеарного пути проявляется двигательными расстройствами только в мышцах языка и нижней части мимической мускулатуры на противоположной стороне. Двустороннее поражение кортико-нуклеарных путей вызывает двусторонние двигательные расстройства в мышечных группах, иннервируемых и другими черепномозговыми нервами.
Кортико-спинальные волокна пирамидной системы на уровне каудальных отделов моста сближаются и на вентральной части продолговатого мозга образуют два макроскопически видимых валика (пирамиды продолговатого мозга). На границе продолговатого и
спинного мозга большая часть волокон (до 80%) переходит на противоположную сторону, а меньшая продолжает идти по своей стороне. В результате в спинном мозге формируется два кортикоспинальных пути. Основным является перекрещенный латеральный кортикоспинальный путь, идущий в боковом канатике. Неперекрещенный прямой кортико-спинальный путь проходит в переднем канатике спинного мозга. На уровне каждого сегмента волокна как прямого, так и латерального кортикоспинальных путей заканчиваются у мотонейронов передних рогов. Прямой кортико-спинальный путь обеспечивает проведение нервного импульса к мотонейронам грудных сегментов, иннервирующих мышцы туловища. Через латеральный кортико-спинальный путь импульсы поступают к мотонейронам всех (в том числе и грудных) сегментов.
Таким образом, мышцы конечностей получают одностороннюю корковую иннервацию от прецентральной извилины противоположного полушария, а мышцы туловища обеспечены двусторонней корковой иннервацией. Этим объясняется тот факт, что при одностороннем поражении кортико-спинального пути в головном мозге (выше перекреста) расстройства произвольных движений возникают в мышцах конечностей на противоположной стороне. Одностороннее поражение кортико-спинального пути в спинном мозге (ниже перекреста) приводит к расстройствам произвольных движений на стороне поражения. Двигательные расстройства в мышцах туловища возможны, но лишь при условии их двустороннего поражения.
Периферический двигательный нейрон образован мотонейронами передних рогов спинного мозга и двигательными ядрами черепно-мозговых 10 нервов, а также их аксонами, достигающими мышцы-исполнители через передние корешки, спинальные нервы, сплетения, периферические или черепные нервы. Каждый мотонейрон иннервирует несколько мышечных волокон, которые образуют «двигательную единицу». Большие α-мотонейроны передают возбуждающий и трофический импульс на быстро сокращающиеся двигательные единицы, а малые α-мотонейроны – на медленно сокращающиеся (тонические) двигательные единицы, γ-мотонейроны посылают нервный импульс на мышечные веретена. Постоянная импульсация от γ-мотонейронов на мышечные веретена является необходимым условием для поддержания мышечного тонуса. Таким образом, периферический двигательный нейрон реализуется через эфферентную часть сегментарных рефлекторных дуг спинальных рефлексов, выполняет тонотропную и трофотропную функции. Центральный двигательный нейрон, заканчиваясь на α-и γ-мотонейронах, регулирует эти сегментарные двигательные функции. Поэтому патология корково-мышечного пути в пределах центрального и периферического двигательного нейронов имеет один общий признак – расстройство произвольных движений. В то же время, двигательные расстройства при поражении центрального и периферического двигательного нейрона существенно отличаются по клинической симптоматике.
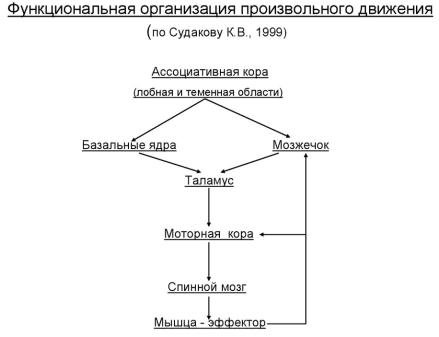
Роль двигательных областей коры, базальных ганглиев и таламуса в организации движений
Важнейшей двигательной областью коры является прецентральная извилина (рис. 66, а). Ее функции были изучены путем раздражения обнаженной поверхности мозга и анализа параличей у больных инсультами. В 50-х годах XX в. американским ученым В.
Пенфилдом были обнаружены интересные закономерности: во-первых, двигательная кора организована по соматотоническому принципу, т. е. каждый ее участок связан с определенной частью тела, во-вторых, области двигательной коры тех частей тела,
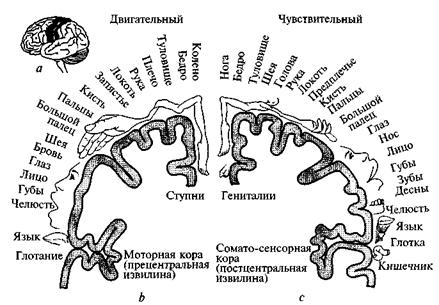
которые осуществляют более разнообразные функции, больше по площади. Наиболее обширными, учитывая пропорции тела, являются зоны, управляющие мышцами кисти руки и мимическими мышцами (см. рис. 66, b). Двигательные и чувствительные зоны коры примыкают друг к другу (см. рис. 66, а) и «чувствительный гомункулюс» почти повторяет карту двигательных зон (см. рис. 66, с). Этот рисунок иногда называют «двигательным гомункулюсом». Предполагается, что определенные типы движений, в которые вовлечены отдельные мышцы, представлены в различных участках двигательной области, причем размеры каждого участка зависят от сложности контролируемых им движений. Например, для участия в речевой функции двигательный центр мозга не только посылает команды к мышцам языка и гортани, но и хранит в памяти последовательность этих команд. Это объясняет, почему рост и развитие полей двигательной области начинаются в раннем детстве и продолжаются вплоть до зрелого возраста. Стимуляция двигательной коры вызывает лишь сокращение отдельных мышц или движение в суставах. Сложные целенаправленные двигательные акты так «запустить» невозможно. По-видимому, двигательная кора является той частью общей структуры регуляции движений, где замысел движения преобразуется в его программу. Фактически двигательная кора — первый компонент структуры регуляции движений, с которого начинается выполнение движения.
Рис. 66. Карта двигательных (b) и сенсорных (с) зон коры головного мозга человека
Нейроны коры, непосредственно связанные с мотонейронами спинного мозга, называются клетками Беца (по имени впервые описавшего их русского анатома XIX в.). Они лежат в глубине двигательной коры и относятся к самым крупным пирамидным нейронам головного мозга. Их аксоны сходятся в толстый пучок нервных волокон, называемый пирамидным трактом. Дойдя до спинного мозга, аксоны клеток Беца перекрещиваются: пучок, идущий от правого полушария, переходит на левую сторону и наоборот. Вот почему регуляция движений левой половины тела контролируется правым (контрлатеральным — противоположным) полушарием, а правой стороны — левым.
Двигательная область коры большого мозга является областью, воспринимающей, анализирующей и синтезирующей раздражения, идущие от скелетно-мышечной системы человека, и участвующей в межанализаторной интеграции. Относительно раннее формирование двигательной коры в онтогенезе, очевидно, определяется ее
функциональной значимостью в обеспечении адекватного поведения детей. Двигательная зона коры обеспечивает осуществление произвольных движений, интегрируя деятельность различных анализаторов всей коры мозга и деятельность всего мозга, благодаря чему осуществляется срочная перестройка путей и форм контактов организма с окружающей средой.
Определенная морфофункциональная зрелость двигательной области коры головного мозга, необходимая для поддержания процессов регуляции движений, отмечается у новорожденных уже с первых дней жизни.
Кроме двигательной области коры головного мозга, в регуляции движений участвуют базальные ганглии, таламус, мозжечок и ствол головного мозга.
Базальные ганглии (скопления нервных клеток, находящихся у основания больших полушарий, формирующиеся на ранних стадиях развития мозга, представляют собой важное подкорковое связующее звено между ассоциативными и двигательными зонами мозга. Базальные ганглии — это четыре образования: полосатое тело (стриатум), бледный шар (паллидум), субталамическое ядро и черная субстанция. Базальные ганглии получают все виды сенсорной информации, и, по-видимому, их функция заключается в «запуске» движений определенного типа — медленных целенаправленных движений конечностей в пространстве. Считается, что на уровне базальных ганглиев имеется готовый набор программ, которые используются в сложных двигательных действиях.
В таламус (структура промежуточного мозга) поступает вся соматосенсорная информация, необходимая для построения любой последовательности движений, и через таламус проходят сигналы от базальных ганглиев и мозжечка к коре.
Мозжечок, имеющий сложную структуру, играет особую роль в нервной регуляции движений, мышечного тонуса и позы.
Следует отметить, что все области коры больших полушарий, в том числе и двигательные, а также другие отделы мозга посылают информацию к мозжечку, к нему же через ассоциативные зоны коры поступают сигналы от периферических органов.
Основное значение мозжечка — дополнение и коррекция деятельности остальных звеньев системы регуляции движений. Экспериментальные данные позволяют выделить следующие функции мозжечка в осуществлении движений: регуляцию позы и мышечного тонуса, коррекцию медленных целенаправленных движений, выполнение последовательности быстрых целенаправленных движений.
В процессе осуществления движений и мозжечок, и базальные ганглии посылают сигналы к двигательной коре через таламус. Обе эти структуры участвуют в выработке программы движений. Таким образом, все эти структуры мозга — двигательная кора, базальные ганглии, таламус и мозжечок исполняют функцию формирования программы целенаправленных движений.
Ствол головного мозга — образование головного мозга, через которое проходят нисходящие пути к спинному мозгу. Эти пути условно можно разделить на два вида — возбуждающие действия мышц сгибателей и возбуждающие действия мышцразгибателей. Они оканчиваются в разных областях спинного мозга. Через ствол мозга проходят и восходящие пути, связывающие между собой структуры ЦНС, которые осуществляют регуляцию движений. Структуры ствола мозга играют важную роль в
регуляции позы, обеспечивающей эффективную реализацию движений (позные компоненты двигательной активности).
Спинной мозг — наиболее древнее образование нервной системы, включенное в структуру регуляции движений.
Нейроны спинного мозга образуют серое вещество, которое имеет на срезе вид буквы «Н». Передние и задние части серого вещества называются передними и задними рогами.
Впередних рогах находятся двигательные нейроны, аксоны которых подходят к мышцам.
Внейронах передних рогов заканчиваются нисходящие пути центральной нервной системы, регулирующие движения. Двигательный нейрон (мотонейрон) и его аксон вместе с мышечными волокнами, которые он контролирует, называют двигательной единицей (ДЕ). Один мотонейрон с помощью разветвлений своего аксона способен контролировать много мышечных волокон. Число волокон, управляемых одним мотонейроном, варьирует в зависимости от того, насколько тонкими должны быть движения мышцы. Например, в глазодвигательных мышцах на каждый нейрон приходится примерно по три мышечных волокна; в мышцах, приводящих в движение бедро, на один нейрон приходится сотня мышечных волокон.
Сила, которую может развить мышца, зависит от числа содержащихся в ней мышечных волокон. У мотонейронов, контролирующих крупные мышцы, такие как бицепсы или мышцы голени, аксоны имеют много разветвлений, для того чтобы передавать импульсы на мышечные волокна, причем веточки аксонов в этом случае гораздо толще, чем у нейронов, управляющих мелкими мышцами пальцев.
В задних рогах находятся нейроны, которые выполняют сенсорные функции и передают сигналы в вышележащие центры, в симметричные структуры противоположной стороны или к передним рогам спинного мозга.
Белое вещество спинного мозга состоит из миелиновых волокон, собранных в пучки. Эти волокна могут быть короткими (связывают нейроны разных сегментов или симметричные нейроны противоположных сторон спинного мозга) или длинными (восходящие — к головному мозгу и нисходящие — от головного мозга к спинному).
В каждой мышце есть чувствительные сенсорные нервы, по которым передается проприоцептивная информация — информация о положении и движении
собственного тела, о напряжении мышцы, о положении сустава, от которых передается информация к чувствительным нервам. Специальные датчики — рецепторы — находятся либо в глубине мышцы, либо в сухожилиях — местах прикрепления мышцы к кости. Эта информация передается либо в спинной мозг, либо в вышележащие нервные центры.
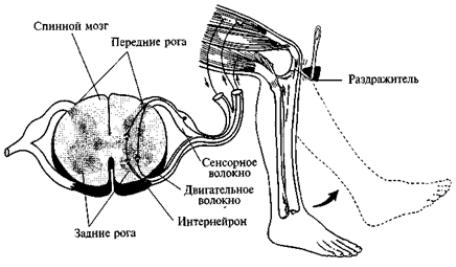
Рис. 67. Коленный
рефлекс Спинальные рефлексы. Особый интерес представляет собственно рефлекторная деятельность спинного мозга (так называемые спинальные
рефлексы) — относительно простые виды реакции, которые осуществляются на уровне спинного мозга без участия вышележащих структур.
Рассмотрим некоторые рефлексы спинного мозга на примере коленного рефлекса (рис. 67) и рефлекса болевого раздражения при уколе пальца иглой (кнопкой) (рис. 68).
В первом случае при ударе молоточком по сухожилию ниже коленной чашки растягивается расположенное выше сухожилие, прикрепленное непосредственно к четырехглавой мышце бедра. В результате активируются находящиеся в этом сухожилии рецепторы, которые по сенсорным волокнам передают возбуждение спинальным мотонейронам, и последние заставляют мышцы бедра сократиться, а ногу — подпрыгнуть. Весь рефлекс совершается очень быстро, обычно меньше чем за секунду.
Другие локальные реакции, которые осуществляются на уровне спинного мозга, связаны, например, с болевыми раздражителями.
При ударе током или случайном уколе рука отдергивается еще до того, как ощущается боль. В этом случае по чувствительным нервам информация передается в спинной мозг, а по двигательным нервам мгновенно передается сигнал к мышцам.
Внутренние системы спинного мозга осуществляют координацию работы мышц сгибателей и разгибателей, позволяя уравновешивать движения рук, ног, тела при выполнении различных по сложности движений.
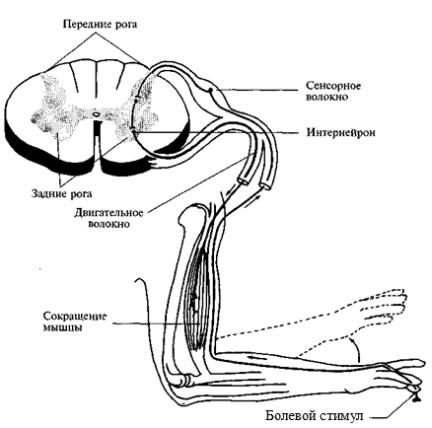
Рис. 68. Рефлекс болевого
раздражения
И все-таки при выполнении большинства движений мышцы сокращаются, т. е. движение реализуется, только если мы этого хотим, если это «не ответ на внешнее раздражение, а решение задачи» (Н.А. Бернштейн). При регуляции произвольных движений в двигательной системе осуществляется последовательная переработка нервных сигналов
— от инициации движения моторной корой до сокращения мышц, контролирующих положение и стабильность суставов, по командам спинальных мотонейронов. Параллельные модифицирующие системы мозжечка, базальных ганглиев ствола мозга обеспечивают координированное и гладкое выполнение двигательной программы, поддержание необходимой позы, эффективность решения двигательной задачи.
20. Автономная (вегетативная) нервная система. Симпатический и парасимпатический отделы. Принципы организации афферентного и эфферентного звена вегетативных рефлексов. Вегетативные ганглии, их функции и механизмы передачи возбуждения.
Вегетативная нервная система (ВНС) представляет часть центральной нервной системы, которая обеспечивает регуляцию деятельности внутренних органов, сосудов и потовых желез, а также трофическую иннервацию скелетной мускулатуры, рецепторов и самой нервной системы. Главная функция ВНС состоит в поддержании постоянства внутренней среды или гомеостаза при различных воздействиях на организм.
На основании структурно-функциональных особенностей различают три отдела вегетативной нервной системы – симпатический, парасимпатический и метасимпатический. Как правило, первые два на органы оказывают противоположные влияния, а третий – обеспечивает саморегуляцию органов.
ВНС отличается от соматической локализацией своих ядер в ЦНС, очаговым выходом волокон из мозга, отсутствием сегментарности их распределения на периферии и малым диаметром волокон. Помимо этого, для ВНС характерно, что ее волокна, направляющиеся из мозга к внутренним органам, обязательно прерываются в периферических вегетативных ганглиях, образуя синапсы на их нейронах. Соответственно, волокна, прерывающиеся в вегетативном ганглии, называются преганглионарными, а выходящие из ганглия – постганглионарными. Аксоны постганглионарных нейронов иннервирует орган-мишень.
Вегетативные ганглии подразделяются на паравертебральные, расположенные по бокам от позвоночника, и превертебральные, лежащие на некотором отдалении от позвоночника. Паравертебральные узлы образуют симпатический ствол. Оба вида ганглиев относятся к симпатическому отделу ВНС.
У парасимпатического отдела ВНС выделяют экстрамуральные (околоорганные) и
интрамуральные (внутриорганные) ганглии.
Центры ВНС расположены в четырех отделах ЦНС – мезенцефальном, бульбарном,
тораколюмбальном и сакральном очагах спинного мозга. Эти центры представлены вегетативными ядрами либо черепных нервов (III, VII, IX, X), либо вегетативными ядрами боковых рогов спинного мозга. Ядра, находящиеся в мезенцефальном, бульбарном и сакральном отделах, образуют пара- 8 симпатическую часть ВНС, а находящиеся в тораколюмбальном отделе – ее симпатическую часть. Дуга вегетативного рефлекса построена по общему принципу, но имеет особенности.
Афферентная часть дуги вегетативного рефлекса существенно не отличается от таковой соматического рефлекса. Одно и то же афферентное волокно может вызвать и вегетативный, и соматический рефлекс. Различаются эти дуги по расположению эфферентных нейронов. У вегетативной рефлекторной дуги они находятся в ганглиях, т.е. вне ЦНС.
Таким образом, вегетативная и соматическая нервные системы действуют содружественно. Однако, они вовлекаются в рефлекторные реакции в разной степени и с разной скоростью.
Все вегетативные рефлексы можно разделить на три группы:
I-я группа - истинные рефлексы – это вегетативные рефлексы, дуга которых состоит из пяти классических звеньев. В зависимости от локализации рецепторов и от того, какой эффектор приходит в деятельное состояние, различают следующие виды истинных вегетативных рефлексов:
1. висцеро-висцеральные – при котором возбуждение начинается и заканчивается во внутренних органах. Например, изменение деятельности желудка
при наполнении тонкой кишки, торможение деятельности сердца при раздражении рецепторов брыжейки (рефлекс Гольца) и др..
2.Висцеро-соматические – изменение соматической деятельности при раздражении рецепторов внутренних органов. Например, сокращение мышц, движение конечностей при сильном раздражении рецепторов ЖКТ.
3.Висцеро-сенсорные – при которых раздражения внутренних органов сопровождаются изменением сенсорной информации. Например, повышение тактильной и болевой