

Ответы на зачёт по НС
.pdfчервя) у больного нарушаются реакции опоры и прыжка. Нарушение реакции опоры проявляется неустойчивостью больного в положении стоя, особенно в позе Ромберга. Нарушение реакции прыжка приводит к тому, что если врач, встав позади больного и подстраховывая его, толкает больного в ту или иную сторону, то больной падает при небольшом толчке (симптом толкания). При поражении мозжечка походка больного обычно изменена в связи с развитием статолокомоторной атаксии. «Мозжечковая» походка во многом напоминает походку пьяного человека, поэтому ее иногда называют «походкой пьяного». Больной из-за неустойчивости идет неуверенно, широко расставляя ноги, при этом его «бросает» из стороны в сторону. А при поражении полушария мозжечка он отклоняется при ходьбе от заданного направления в сторону патологического очага. Особенно отчетлива неустойчивость при поворотах. Если атаксия оказывается резко выраженной, то больные полностью теряют способность владеть своим телом и не могут не только стоять и ходить, но даже сидеть.
Преимущественное поражение полушарий мозжечка ведет к расстройству его противоинерционных влияний, в частности к возникновению кинетической атаксии. Она проявляется неловкостью движений и особенно выражена при движениях, требующих точности. Для выявления кинетической атаксии проводятся пробы на координацию движений. Далее приводится описание некоторых из них.
Мозжечок (cerebellum, малый мозг) — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций.
Особенности морфофункциональной организации и связи мозжечка.Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка:
1)кора мозжечка построена достаточно однотипно, имеет стереотипные связи, что создает условия для быстрой обработки информации;
2)основной нейронный элемент коры — клетка Пуркинье, имеет большое количество входов и формирует единственный аксонный выход из мозжечка, коллатерали которого заканчиваются на ядерных его структурах;
3)на клетки Пуркинье проецируются практически все виды сенсорных раздражений: проприоцептивные, кожные, зрительные, слуховые, вестибулярные и др.;
4)выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом.
14. Ретикулярная фармация. Ее стркуктурнофункциональная организация, особенности свойств ее нейронов. Роль ретикулярной фармации в регуляции двигательной активности. Нисходящии влияния рф (тормозящие и облегчающие). Восходящие влияния ретикулярной фармации. Участие рф в интегративной деятельности цнс.
Ретикулярная формация — это совокупность нервных клеток и их отростков, расположенных в покрышке всех уровней ствола между ядрами черепных нервов, оливами, проходящими здесь афферентными и эфферентными проводящими путями. К ретикулярной формации иногда от носят и некоторые медиальные структуры промежуточного мозга, в том числе медиальные ядра таламуса.

Строение ретикулярной формации Ретикулярная формация - совокупность нейронов отростки которых образуют своеобразную сеть в пределах центральной нервной системы.
В ретикулярной формации ствола мозга различают 2 отдела: растральный - ретикулярная формация на уровне промежуточного мозга;
каудальный - ретикулярная формация продолговатого мозга, моста и среднего мозга.
Изучены 48 пар ядер ретикулярной формации.
Особенности нейронов ретикулярной формации: постоянная спонтанная электрическая активность - обеспечивается гуморальным влиянием и влиянием вышележащих отделов центральной нервной системы. Эта активность не имеет рефлекторного происхождения;
явление конвергенции - к ретикулярной формации идут импульсы по коллатералям различных проводящих путей. Сходясь к телам одних и тех же нейронов импульсы теряют свою специфичность; импульсы, поступая к нейронам ретикулярной формации, изменяют ее функциональную активность - если нейроны обладают выраженной электрической активностью, то под влиянием афферентных импульсов электрическая активность уменьшается и наоборот, т. е. модулируется активность нейронов ретикулярной формации; у нейронов ретикулярной формации низкий порог раздражения и, как следствие, высокая возбудимость; у нейронов ретикулярной формации высокая чувствительность к действию гуморальных факторов: биологически активных веществ, гормонов (адреналина), избытку СО2, недостатку О2 и т. д.; в состав ретикулярной формации входят нейроны с различными медиаторами: адренэргические, холин-, серотонин-, дофаминэргические.
Роль рф в двигательной активности. В целом ретикулярная формация выполняет следующие функции: Выбирает тип поведения всего организма в зависимости от конкретной обстановки. Оказывает облегчающее или тормозящее влияние на сгибательные и разгибательные рефлексы, рефлексы поддержания позы, физическую двигательную активность.
Нисходщяие влияния рф ( торможение, облегчение)
Восходящее влияние Рф
15. Таламус. Его структурнофункциональная организация. Функции специфических, ассоциативных, и неспецифических ядер. Роль в регуляции двигательной активности и в интегративной деятельности мозга. Участие ядер таламуса в формировании болевых ощущений. Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга
Участие ядер таламуса в формировании болевой чувствительности
16. Лимбическая система и ее роль в формировании мотиваций, эмоций, формирования памяти. Участие лимбических структур в регулиции соматических и вегетативных функций и интегративной деятельности ЦНС.
Лимбическая система — совокупность нервных структур и их связей, расположенных в медиобазальной части больших полушарий, участвующих в управлении вегетативными функциями и эмоциональным, инстинктивным поведением, а также оказывающих влияние на смену фаз сна и бодрствования. К лимбической системе относится наиболее

древняя часть коры головного мозга, расположенная на внутренней стороне больших полушарий
17. Базальные ганглии, их структурно функциональная организация. Функции полосатого тела, субталомического ядра, бледного шара. Роль в формировании тонуса и сложных двигательных актов , в организации и реализации двигательных программ, в интегративной функции ЦНС.
Базальные ганглии, или подкорковые ядра, относятся к структурам переднего мозга и включают в себя полосатое тело, или неостриатум (хвостатое ядро и скорлупа), палеостриатум (бледный шар) и ограду. Эта структура мозга играет главную роль в процессе перехода от замысла (фазы подготовки) движения к выбранной программе действия (фазе выполнения движения).
Базальные ганглии включают в себя следующие компоненты: бледный шар (pallidum) - наиболее древнее образование стриопаллидарной системы;
неостриатум - в его состав входят полосатое тело и скорлупа (putamen);
ограда (claustrum) - самое новое образование.
Связи базальных ганглиев: внутри, между базальными ганглиями. За их счет компоненты базальных ганглиев тесно взаимодействуют и образуют единую стриопаллидарную систему; связь с образованиями среднего мозга. Они носят двусторонний характер за счет дофаминэргических нейронов. За счет этих связей стриопаллидарная система тормозит активность красных ядер и черной субстанции, которые регулируют мышечный тонус; связь с образованиями промежуточного мозга (таламусом и гипоталамусом); с лимбической системой; с корой головного мозга.
Функции полосатого тела Полосатое тело состоит из более крупных нейронов с длинными отростками, которые выходят за пределы стриопаллидарной системы. Полосатое тело регулирует мышечный тонус, уменьшая его; участвует в регуляции работы внутренних органов; в осуществлении различных поведенческих реакций (пищедобывающее поведение); участвует в формировании условных рефлексов.
При разрушении полосатого тела происходит: гипертонус скелетных мышц, нарушение сложных двигательных реакций и пищедобывающего поведения; тормозится формирование условных рефлексов. Это проявляется: хореей (пляска святого Витта) - навязчивые двигательные реакции, нет содружественных и вспомогательных двигательных реакций; нарушение поведенческих реакций; снижение интеллекта.
Функции ограды Ограда. Функции ограды: участвует в регуляции мышечного тонуса; участвует в эмоциональных реакциях; участвует в формировании условных рефлексов.
Функции бледного шара Бледный шар состоит в основном из мелких нейронов (на 95%) и их отростки, как правило, коротки. Имеет тесные связи с образованиями среднего и промежуточного мозга.
Функции:
регулирует мышечный тонус, участвует в регуляции двигательной активности;
участвует в эмоциональных реакциях за счет влияния на мимическую мускулатуру;
участвует в интегративной деятельности внутренних органов, способствует объединению функции внутренних органов и мышечной системы.
При раздражении бледного шара наблюдается резкое снижение мышечного тонуса, замедление движений, нарушение координации движений, деятельности внутренних органов (сердечно-сосудистой и пищеварительной систем). При сосудистых поражениях бледного шара развиваетсяболезнь Паркинсона - нарушается мышечный тонус (в одних группах мышц повышается, в других - понижается); нарушаются сложные двигательные функции, мимическая активность (лицо-маска); возникает мышечная дрожь (тремор), который усиливается в состоянии покоя.
Роль базальных ядер в осуществлении сложных двигательных актов заключается в том, что они обусловливают миостатические реакции, оптимальное перераспределение мышечного тонуса (благодаря модулирующим влияниям на нижележащие структуры центральной нервной системы, определяющие регуляцию движений).
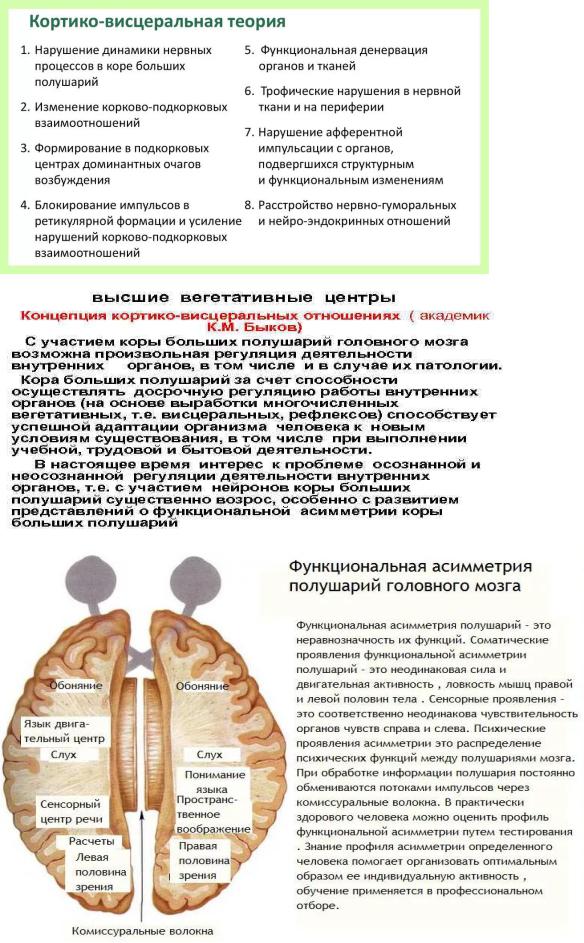
18. Кора больших полушарий, ее структурно-функциональная организация. Сенсорные, моторные и ассоциативные области коры. Современные представления о локализации функций в коре. Пластичность коры, корково-подкорковые и кортиковисцеральные взаимоотношения. Функциональная асимметрия коры больших полушарий у человека. Кора больших полушарий (неокортекс) представлена скоплением нервных клеток, имеет многослойное строение (состоит из 6 слоев), множество борозд и извилин, за счет которых создается большая ее поверхность (1500– 2200 см2). Новая кора занимает около 72 % всей площади коры и около 40 % массы головного мозга. В коре имеется 14 млрд нейронов, количество глиальных клеток примерно в 10 раз больше.
К основным бороздам коры относят боковую, центральную, теменно-затылочную, поясную и коллатеральную мозговые борозды. Они делят кору больших полушарий на доли. Различают шесть основных долей: лобную, теменную, височную, затылочную, краевую и островок. Каждая доля вторичными бороздами делится на извилины. По классификации К. Бродмана, кора разделена на 52 поля. С полями коры больших полушарий связаны ее регуляторные влияния на функции организма.
Кора головного мозга является наиболее молодой в филогенетическом отношении структурой мозга, осуществляющей у человека высшие интегративные функции контроля и регуляции соматических, висцеральных и психических функций.
В коре различают шесть горизонтальных слоев.
I. Молекулярный слой образован дендритами пирамидных клеток, формирующих сплетение, расположенное параллельно поверхности. На дендритах образуют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
II. Наружный зернистый слой представлен звездчатыми клетками и частично малыми пирамидными клетками. Волокна этого слоя образуют кортикокортикальные связи.
III. Наружный пирамидный слой состоит из пирамидных клеток средней величины. Аксоны этих клеток образуют также кортикокортикальные ассоциативные связи.
IV. Внутренний зернистый слой образован звездчатыми клетками. На этих клетках проецируются афферентные волокна нейронов специфических ядер таламуса, получающих информации от рецепторов сенсорной системы.
V. Внутренний пирамидный слой образован средними и крупными пирамидными клетками, аксоны которых образуют кортикоспинальные пути.
VI. Слой полиморфных клеток образован веретенообразными клетками, аксоны которых образуют кортикоталамические пути.
Выраженность отдельных слоев в различных областях коры не одинакова. По классификации К. Бродмана, основанной на цитоархитектонических признаках (формы и расположения нейронов), кора разделена на 52 поля. С полями коры больших полушарий связаны ее регуляторные влияния на функции организма (рис. 3.12).
Электрическая активность коры больших полушарий характеризуется специфическими ритмами определенной частоты и амплитуды, которые обозначаются термином электроэнцефалограмма (ЭЭГ).
Для отведения ЭЭГ существуют два способа: биполярный и монополярный. При биполярном отведении регистрируется разность потенциалов между двумя активными электродами. При монополярном отведении регистрируется разность потенциалов между активной точкой на поверхности головы и индифферентной точкой головы или лица, где электрические процессы минимальные и их можно условно принять за нуль (обычно это сосцевидный отросток черепа или мочка уха). Выбор того или иного варианта записи зависит от целей исследования. В исследовательской практике шире используется монополярный вариант регистрации, поскольку он позволяет изучать изолированный вклад работы той или иной зоны мозга в изучаемый процесс.
В ЭЭГ человека выделяют следующие ритмы (рис. 3.13).
Альфа-ритм. Это наиболее часто встречающийся ритм, который состоит из волн правильной, почти синусоидальной формы с частотой 8–13 Гц и с амплитудой 50–100 мкВ. Регистрируется альфа-ритм преимущественно в затылочной области коры. Наблюдается он в состоянии покоя и при отсутствии внешних раздражений. Этот ритм слабо выражен или отсутствует у слепых. При переходе человека от состояния покоя к деятельности (восприятие зрительных или акустических сигналов, умственная работа и т. д.) альфа-ритм десинхронизируется, т. е. заменяется низкоамплитудным, высокочастотным бета-ритмом. Синонимами этого термина могут быть реакция активации, реакция открывания глаз или реакция пробуждения.
Выделяют также некоторые эквиваленты альфа-ритма, имеющие другую локализацию и чувствительные к другим видам модальности. В области роландовой борозды регистрируется мю-ритм (роландический), отвечающий блокадой на проприорецептивные раздражения. В височной коре регистрируется тау-ритм, который подавляется звуковыми стимулами. Каппа-ритм имеет также одну частотную характеристику с альфа-ритмом. Он регистрируется в височной области при подавлении альфа-ритма в других областях в процессе умственной деятельности.
Бета-ритм имеет частоту 18–30 Гц и амплитуду колебаний около 2–20 мкВ. Этот ритм регистрируется в прецентральной и фронтальной коре, а также и в других областях коры при различных видах интенсивной деятельности.
Гамма-ритм имеет частоты от 30 до 120–170 Гц, а по некоторым данным – до 500 Гц, при амплитуде не превышающей 15 мкВ. Этот ритм можно наблюдать в прецентральной, фронтальной, височной, теменной и специфических зонах коры. Наблюдается при решении задач, требующих максимального сосредоточения внимания.
Тета-ритм включает колебания с частотой 4–8 Гц, амплитудой 5–100 мкВ и чаще наблюдается во фронтальных зонах коры, выражен в гиппокампе. Связан этот ритм с поисковым поведением, усиливается при эмоциональном напряжении.
Дельта-ритм состоит из самых медленных волн с частотой 0,5–4,0 Гц, амплитудой 20– 200 мкВ. Возникает при естественном и наркотическом сне, а также при некоторых патологических состояниях коры.
Кора больших полушарий • Кора больших полушарий головного мозга или кора головного мозга (лат. cortex cerebri) — структура головного мозга, слой серого вещества толщиной 1,3—4,5 мм, расположенный по периферии полушарий большого мозга, и покрывающий их.
•Всю кору полушарий принято разделять на 4 типа: древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора.
•В коре больших полушарий различают три области: сенсорные, моторные и ассоциативные.
1. Сенсорные (чувствительные) зоны локализованы в различных областях коры: зрительная -в затылочной области полушарий, слуховая - в височной, зона вкусовых ощущений - в нижней части теменных областей, соматосенсорная - в области задней центральной извилины. Сенсорные области коры больших полушарий выделяют отдельные признаки сигнала. Так, если это в области зрительной проекции, то анализируются место объекта в поле зрения, направление движения, контур, цвет, контраст. Если в области слуховой проекции, то анализируются признаки звука.
2. Зоны, раздражение которых вызывает двигательную реакцию, называют моторными или двигательными. Движения возникают при раздражении коры в области предцентральной извилины. Величина корковой двигательной зоны пропорциональна не масса мышц, п точности движений. Особенно велика зона, управляющая движениями кисти рук, языком, мимической мускулатурой. Моторная кора имеет двусторонние внутрикорковые связи со всеми сенсорными областями, что обеспечивает их взаимодействие.
3. Ассоциативные области коры не имеют прямых афферентных и эфферентных связей с периферией, нейроны этих областей не связаны не с органами чувств, ни с мышцами. Они осуществляют связь между различными областями коры, интегрируя, объединяя все поступающие в кору импульсы в целостные акты научения (чтение, речь, письмо), логического мышления, памяти и обеспечивая возможность целесообразной реакции поведения.

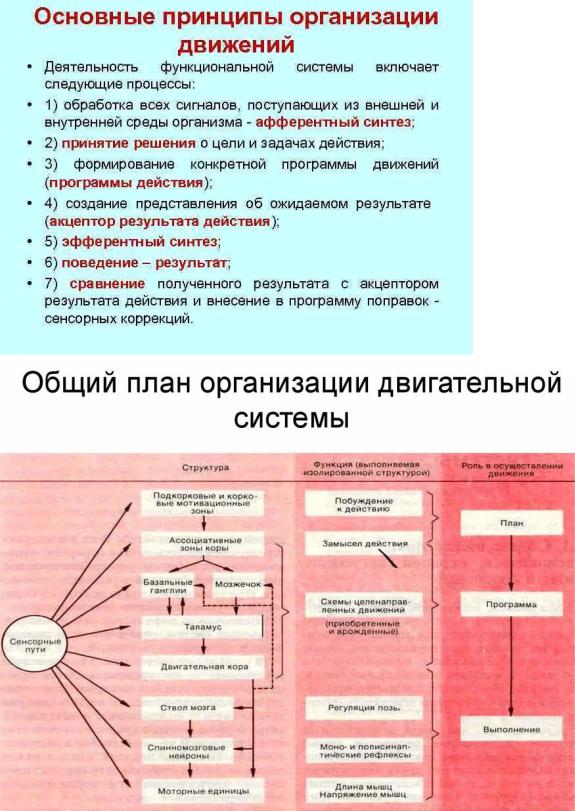
19. Общая концепция организации движений и целевых функций двигательной системы. Роль спинальных, шейных и вестибулярных рефлексов. Произвольные движения: функциональные блоки, их организация. Роль коры, мозжечка, базальных ганглиев, лимбической системы, таламуса, ствола, спинного мозга в организации и осуществлении двигательных актов.
Рефлексы продолговатого мозга Продолговатый мозг является жизненно важным отделом нервной системы, где замыкаются дуги целого ряда соматических и вегетативных
рефлексов. При участии ядер продолговатого мозга осуществляются цепные рефлексы, связанные с жеванием и проглатыванием пищи. Так, например, при акте жевания координированная деятельность жевательных мышц, языка, щек, нёба и дна полости рта обеспечивается функцией моторных ядер тройничного и подъязычного
нервов. Когда пищевой комок касается нёба, основания языка или задней стенки глотки, возникает цепной рефлекс глотания. От рецепторов задней трети языка, слизистой оболочки глотки возбуждение по чувствительным волокнам языкоглоточного нерва поступает в продолговатый мозг к центру глотательного рефлекса. Из этого центра запрограммированные в нервной системе моторные команды по двигательным волокнам подъязычных, тройничных, языкоглоточных и блуждающих нервов идут к мышцам полости рта, глотки, гортани и пищевода. В результате координированной деятельности этих мышц совершается акт глотания с одновременным перекрытием входов в носоглотку и гортань.
С функцией пищеварительного тракта связаны многие вегетативные рефлексы заднего мозга. К их числу относится, например рефлекторная регуляция секреции слюнных желез, которая осуществляется с участием ядер лицевого и языкоглоточного нервов, содержащих парасимпатические нейроны. Вегетативное, парасимпатическое ядро блуждающего нерва включено в систему рефлекторной регуляции дыхания, деятельности сердца и тонуса сосудов.
Целая группа рефлексов заднего мозга направлена на поддержание позного тонуса. В зависимости от источника рецепторных воздействий тонические рефлексы подразделяются на шейные и вестибулярные, или лабиринтные.
Шейные тонические рефлексы запускаются при возбуждении проприоцепторов мышц шеи. Впервые они описаны голландским физиологом Р. Магнусом на кошках с перерезанным на уровне шатра мозжечка стволом мозга (см. разд. 3.5.2). В чистом виде шейные .рефлексы можно наблюдать при разрушении вестибулярного аппарата, дающего дополнительную информацию о положении головы в пространстве. Как показали опыты Р. Магнуса, запрокидывание головы кошки назад в сагиттальной плоскости вызывает возрастание тонуса мышц—разгибателей передних конечностей и одновременно ослабление экстензорного тонуса задних. Напротив, наклон головы вниз вызывает обратный процесс, когда тонус разгибателей передних конечностей снижается, а задних
— возрастает. Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова. Это так называемый рефлекс вращения по Магнусу.
Эффекторным звеном шейных рефлексов могут быть не только мышцы конечностей, но и глазная мускулатура. Рефлекторные влияния на глазные мышцы обеспечивают компенсаторную установку глазных яблок при изменении положения головы. Любое отклонение головы вызывает движение глазных яблок в противоположном направлении, и таким образом рефлекторно сохраняется правильная зрительная ориентация животного.
Все шейные рефлексы являются полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в соответствующие центры продолговатого мозга, а оттуда нисходящие команды поступают или к мотонейронам спинного мозга, или к двигательному ядру отводящего нерва и производят определенное влияние на позный тонус и на глазную мускулатуру.