

3. Положение микробов среди живых существ!!!!!!!!!!!!.
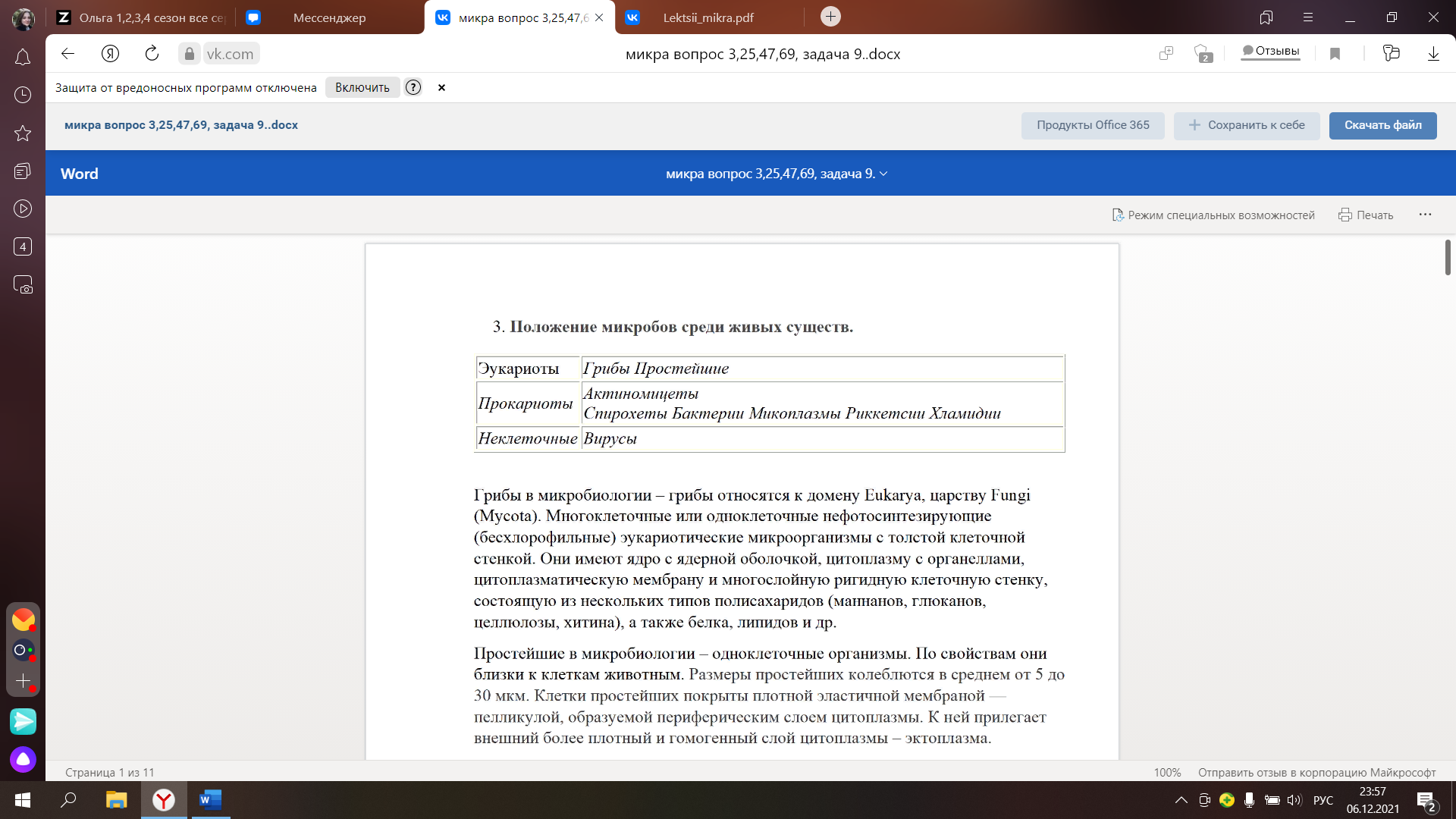
Грибы в микробио – грибы относятся к домену Eukarya, царству Fungi (Mycota). Многоклет или одноклет нефотосинтезирующие (бесхлорофильные) эукариотические микроорг с толстой клет стенкой. Они имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клет стенку, состоящую из нескольких типов полисахаридов (маннанов, глюканов, целлюлозы, хитина), а также белка, липидов и др.
Простейшие в микробио – одноклет орг. По св-вам они близки к кл животным. Размеры простейших колеблются в среднем от 5 до 30 мкм. Кл простейших покрыты плотной эластичной мембраной — пелликулой, образуемой периферическим слоем цитоплазмы. К ней прилегает внешний более плотный и гомогенный слой цитоплазмы – эктоплазма.
Актиномицеты. Порядок бак, имеющих способность к формированию на некот стадиях развития ветвящегося мицелия, кот проявл у них в оптимальных для существования условиях. Некот исслед, подчёркивая бактериал природу актиномицетов, называют их аналог грибного мицелия тонкими нитями; их диаметр 0,4-1,5 мкм. Актиномицеты имеют кислотоустойчивый тип клет стенки, кот окрашивается по Граму как грам+, однако по структуре ближе к грам-. Характ высоким содержанием ГЦ пар в ДНК
Спирохеты. Спирохетозы – это гр инфекц заб, вызываемых спиралевидными бак, характ общей интоксикацией и цикличностью развития. Длинные, тонкие, спирально-изогнутые микроорг, размеры 0,1-0,3х5-250 мкм. Центр структурой явл протоплазматический цилиндр, в кот содержатся цитоплазма, нуклеоид, рибосомы и ферменты, окруж цитоплазматической мембраной. Протоплазматич цилиндр имеет постоянную спиралевидную форму, благодаря пептидогликану, образуя первич завитки. Их число, тип, шаг, угол наклона варьируют у разных видов. В результате изгиба всего тела образуются вторич завитки. Вокруг спиралевидного протоплазматического цилиндра располаг периплазматические жгутики (эндожгутики), 1 конец их прикреплен к 1му из полюсов протоплазматического цилиндра, а др – к последнему примерно посередине кл, кол-во жгутиков от 2 до 100 (1 половина прикреплена к 1му полюсу, а 2ая – к др). Периплазматические жгутики спирально обвивают протоплазматический цилиндр, образуя осевую нить, поверх кот располаг многослойная наруж клет оболочка спирохеты, т е жгутик расположен между ней и цилиндром (поэтому и называется эндожгутиком). C помощью жгутиков спирохеты активно передвигаются (жгутики сокращаются вокруг своей оси, совершая поступательные и сгибательные движения). Спор и капсул не образуют. При неблагоприятных условиях образуют цисты (сворачиваются в клубок, выделяя слизь и покрываясь муциновой оболочкой), могут превращаться в L-формы.
Бак (гр.bakterion-палочка)- простые одноклет микроскопические орг, принадлежащие к прокариотам.
4. Основные формы бак. Морфология, ультраструктура, химич состав бактериал кл. Основные отличия прокариотов от эукариотов. Протопласты, сферопласты, l-формы бак.
Среди основных морфологических форм бактерий различают:
шаровидные (кокковые), кот по характеру взаиморасполож делятся на: Микрококки-отдельное изолированное располож. Диплококки-сцепленные попарно, т.к. кл после деления не расходятся. Стрептококки-сцепленные в цепочку, вследствие деления кл в 1ой плоскости и сохранения связи между ними в месте деления. Сарцины-сцепленные в пакеты по 8, 12, 16, т.к. они образуются при делении клетки в 3х взаимно перпендикулярных плоскостях. Стафилококки-сцепленные беспорядочно в виде виноградной грозди, т.к. кл делятся в разных плоскостях.
палочковидные, кот различаются по форме – правильная (энтеробак, псевдомонады), неправ (коринебак), размерам – мелкие (бруцеллы, бордетеллы), сред (бактероиды, кишеч палочка), крупные (бациллы, клостридии), форме концов – обрубленные (бациллы), закругленные (сальмонеллы, псевдомонады), заостренные (фузобак), утолщенные (коринебак), а по характеру взаимораспол все палочки делятся на: расположенные по одиночке, диплобактерии и диплобациллы (сцепленные попарно), стрептобактерии и стрептобациллы (сцепленные в цепочку), слегка изогнутые (вибрионы).
извитые формы. По характеру и кол-ву завитков они делятся на спириллы (1 или несколько завитков), и спирохеты, кот делятся на лептоспиры (завитки с загнутыми крючкообразными концами – S-образная форма), боррелии (4-12 неправильных завитков) и трепонемы (14-17 равномерных мелких завитков).
Химич состав бактериал кл: Вода 70-80% Сухой остаток 15-30% от массы микробной кл Белки 52% сухой массы Нуклеин к-ты 19% сух м Липиды 9% сух м Углеводы 17% сух м Минеральные в-ва 3% сух м
Ультраструктура: Структура бактериал кл м б разделена на такие компоненты, как обязательные и необязательные. К обязат компонентам относится: Оболочка, кот состоит из клет стенки и ЦПМ.
Протоплазма, кот состоит из цитоплазмы, и расположенными в ней нуклеоидом и рибосомами.
Рибосомы бак имеют коэффициент седиментации 70S в отличие от 80S-рибосом, характерных для эукариотических кл. Поэтому некот а/б, связываясь с рибосомами бак, подавляют синтез бактериал белка, не влияя на синтез белка эукариотич кл. Рибосомы бак могут диссоциировать на 2субъединицы — 50S и 30S. Нуклеоид — эквивалент ядра у бак. Он расположен в центр зоне бак в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка.
Необязательные компоненты: Включения (в виде гранул гликогена, полисахаридов, оксимасляной кислоты и полифосфатов (волютин)) Капсула (Слиз структура толщиной более 0,2 мкм, прочно связанная с клет стенкой бак и имеющая четко очерченные внешние границы) Микрокапсула (Слиз образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии) Слизь (Мукоидные экзополисахариды, не имеющие четких внеш границ. Слизь растворима в воде) Жгутики (Тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама кл. Жгутики состоят из белка флагеллина являющ антигеном, так называемый Н-антиген) Пили (Фимбрии, ворсинки — нитевидные образования, более тонкие и короткие, чем жгутики. Пили отходят в большом кол-ве (за искл половых) от поверхности кл и состоят из белка пилина. Они обладают антигенной активностью) Споры (своеобразная форма покоящихся бак с грам+ типом строения клет стенки. Споры образуются при неблагоприятных условиях существования бактерий)
Основные отличия прокариот от эукариот:
Отсутствие истинного дифференцированного ядра.
Отсутствие развитой ЭПС, аппарата Гольджи.
Отсутствие митохондрий, хлоропластов, лизосом.
Неспособность к эндоцитозу.
Как правило прокариоты значительно меньше эукариот по размеру.
В отличии от эукариот, прокариоты имеют мезосомы.
Кольцевая молекула ДНК у прокариот.
Рибосомы 70S.
Клеточная стенка из муреина.
Протопласты — бактерии, полностью лишенные клеточной стенки;
Сферопласты — бак с частично сохранивш клет стенкой. После удаления ингибитора клет стенки такие измененные бак могут реверсировать, т.е. приобретать полноценную клет стенку и восстанавливать исходную форму.
L-формамы- бак сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием а/б или др ф-ров и способные размножаться. Они предст собой осмотически чувствительные, шаровидные, колбовидные кл различной величины. Некот L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать.
5. Структура бактериал кл. Перечислите постоянные и непостоянные структуры, назовите основные отличия прокариотов от эукариотов. Укажите химич строение, биологическую роль и методы выявления клет стенки, цитоплазматической мембраны, мезосом, рибосом, никлеоида, цитоплазмы.
Постоянные структуры: оболочка, состоящая из клет стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и наследственного аппарата — аналога ядра, называемого нуклеоидом.
Дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Основные отличия прокариотов от эукариотов:
Отличительный признак |
Эукариотич кл |
Прокариотич кл |
Наличие истинного ядра, отделенного от цитоплазмы ядерной мембраной, в кот присутствуют ядрышко и связанные с молекулой ДНК белки-гистоны |
+ |
Истинное ядро отсут, вместо него присут нуклеоид с гаплоидным набором генов |
Наличие в цитоплазме вторич мембранных образований (митохондрии, аппарат Гольджи, эндоплазматический ретикулум) |
+ |
- |
Присутствие сторолов в цитоплазматической мембране |
+ |
- (искл микоплазмы) |
Рибосомы |
Типа 80S |
Типа 70S |
Наличие в клет стенке пептидогликана |
+ |
- |
В зависимости от строения клет стенки прокариоты, относящиеся к эубактериям, делятся на 2 большие гр: грам+ и грам- бактерии.
Основа -пептидогликан, обеспечивающий ригидность и эластичность КС.
Структура: N-ацетилглюкозамина и N-ацетилмурамовой к-ты, соединенных между собой посредством гликозидных связей.
К каждому остатку N-ацетилмурамовой к-ты присоединен короткий пептид из 4-5 аминок-т.
2 особенности пептидного хвоста заслуживают внимания: наличие аминок-т в D-форме (неприродная конфигурация) и высокое содержание аминок-т с 2мя аминогр. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей.
У грам+ эубак он составляет основную массу в-ва клет стенки (от 40 до 90%), у грам- — содержание пептидогликана значительно меньше (1—10%).
Особенности строения КС Гр+ Клет стенка грам+ бактерий достаточно плотно прилегает к ЦПМ.
Пептидные сшивки в пептидогликане обеспечивают его 3хмерную пространств организацию.
Многослойный пептидогликан пронизывают тейхоевые к-ты – полифосфатные соединения на основе рибитола или глицерина.
Тейхоевые к-ты ковалентно могут соединяться с N-ацетилмурамовой к-той (собственно тейхоевые или стеночные) или с гликолипидом ЦПМ (липотейхоевые).
Свободные гидроксилы фосфорной к-ты придают тейхоевой к-те св-ва полианиона, определяющего поверхностный заряд клетки.
Особенности строения КС Гр- Пептидогликан образует только тонкий внутр слой клет ст, неплотно прилегающий к ЦПМ.
У бол-ва видов пептидогликан образует 1- или 2хслойную структуру, характ весьма редкими поперечными связями между гетерополимерными цепями.
Снаружи от пептидогликана располагается дополнит слой клет стенки-наруж мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (ЛПС).
ЛПС сложного молекулярного строения, занимает около 30—40% поверхности наруж мембраны и является ее важнейшим компонентом.
Биологическая роль клеточной стенки:
Поддержание формы клетки
Защита от действия механических и осмотических действий внешней среды
Участие в регуляции роста и деления клетки
Связь с внешней средой через каналы и поры
Определяет антигенную характеристику бактерий
Содержит рецептуры для бактериофагов
Методом выявл клет стенки явл метод Пешкова (КС –красная; цитоплазма –розовая), по Грамму-тип стоения клеточной стенки.
Этапы окраски: 1.мазок протравливать в 10% растворе танина 6-8 мин
2.промыть водой
3.окрашивать водным раствором фуксина 30-60 сек
4.высушить
Сущность метода: танин уплотняет клет стенку бак, и большая часть фуксина задерживается в ней
Цель: выявление клеточной стенки. КС - красная, цитоплазма -розовая.
Химическое строение цитоплазматической мембраны: Состоит ЦПМ из липидов (15–45%), белков (45–60%) и незначительного кол-ва углеводов (около 10%).
Липиды представлены фосфолипидами – до 30% сухой массы самой мембраны. Преобладают фосфатидилглицерин и дифосфатидилглицерин. Кроме того, обнаружены гликолипиды, каротиноиды, хиноны. В составе липидов присутствуют нетипичные жирные к-ты (ненасыщенные и мононасыщенные), циклопропановые и разветвленные жирные к-ты. Набор жирных к-т и состоящих из них липидов для прокариот является видоспецифичным признаком.
Белки составляют половину и более сухой массы мембран. Их насчитывается более 20 типов. Они подразделяются на интегральные (погружены в гидрофобную обл мембраны) и периферические (локализованы на поверхности гидрофильного слоя и часто прикреплены к интегральным белкам).
Углеводы в мембране взаимосвязаны с белками и липидами. Они обычно локализованы только на наружной поверхности и выполняют функции рецепторов опознавания факторов внешней среды.
Биологическая роль цитоплазматической мембраны: Восприятие всей химич инф-ции из внешней среды
Осмотический барьер
Регуляция роста и клеточного деления
Участие в транспорте веществ
Связь со жгутиками
Участие в синтезе компонентов клеточной стенки, образование мезосом.
Метод выявления цитоплазматической мембраны-электронная микроскопия.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов предст собой 3хслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым — промежуточным). По структуре она похожа на плазмолемму кл животных и состоит из двойного слоя липидов, глав обр фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некот из них явл пермеазами, участвующ в транспорте в-в. В отличие от эукариотич кл, в цитоплазматической мембране бактериал кл отсутст стеролы (за исключением микоплазм).
Химич строение мезосом Мезосомы — складчатые структуры, представл собой впячивания плазматич мембраны кл. Имеют такой же состав, что и цитоплазматическая мембрана. Во вр клет деления мезосомы, по-видимому, ассоциируются с ДНК, что обеспечивает разделение 2х дочерних молекул ДНК после репликации и способствует образованию перегородки между дочерними кл.
Биологическая роль мезосом:
аналог митохондрий, обеспечивают синтез АТФ и процессы дыхания
участвуют в делении клетки
участвуют в спорообразовании
Метод определения мезосом: электронная микроскопия.
Химич строение рибосом: Рибосомы бак – сложные глобулярные образования, состоящие из различных молекул РНК и многих связанных с ними белков; все образование функционирует как локус синтеза белков. Рибосомы бак имеют размер около 20 нм и состоят из 2х субъединиц с коэффициент седиментации 50 S и 30 S. Перед началом синтеза белка происходит объединение этих субъединиц в 1 – 70S, в отличие от 80S-рибосом, характ для эукариотич кл. В отличие от эукариотич кл рибосомы бак не объединены в эндоплазматическую сеть. В состав рибосом входят рибосомные РНК (рРНК) – консервативные элементы бак («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК – в состав большой субъединицы рибосом. Изучение 16S рРНК явл основой геносистематики, позволяя оценить степень родства организмов.
Биологич роль рибосом: отвечают за биосинтез белка. Метод обнаружения: электрон микроскопия.
Химическое строение нуклеотида: Нуклеоид - двунитевая молекула ДНК, замкнута в кольцо, лишена мембраны. Нуклеоид бак, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У бол-ва бак содержится 1 хромосома, представленная замкнутой в кольцо молекулой ДНК. Но у некот бак имеются 2 хромосомы кольцевой формы (V. cholerae) и линейные хромосомы. Биологич роль нуклеоида: Носитель генетич инф-ции, расположен в центр зоне бак (1 хромосома) Метод обнаружения нуклеоида: световая микроскопия, электрон микроскоп
Нуклеоид выявл в световом микроскопе после окраски специфическими для ДНК Морфология и классификация микробов методами: по Фельгену или по Романовскому—Гимзе.
На электронограммах ультратонких срезов бак нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной опред участками с цитоплазмаческой мембраной или мезосомой, участвующими в репликации хромосомы.
Химич строение цитоплазмы: Цитоплазма ограничена цитоплазматической мембраной. Она предст собой полужидкую коллоидную массу гранулярной структуры, на 70–80% состоящую из воды и заполняющую внутреннюю полость клетки. Цитоплазма состоит из двух фракций:
цитозоль – имеет гомогенную структуру, содержит набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций;
2ая – представлена структурными элементами, к кот относятся рибосомы, нуклеоид, мембранные структуры, внутриплазматические включения.
Биологическая роль: Обеспечивает функционирование всех органелл.
Создание оптимальных условий для поддержания жизнедеятельности рибосом.
Транспортировка веществ внутри организма.
Объединение внутренней структуры организма, обеспечение необходимого химического взаимодействия между включениями. Метод обнаружения: электронная микроскопия.
6. Структура бактериал кл. Перечислите постоянные и непостоянные структуры, назовите основные отличия прокариотов от эукариотов. (СМ В 5 ВОПРОСЕ!!!!!!!!!!!) Укажите химич строение, биологич роль и методы выявления капсулы, спор, жгутиков, пилей, включений
Капсула — слиз структура толщиной более 0,2 мкм, прочно связанная с клет стенкой бак. выявляется при спец методах окраски мазка по Бурри— Гинсу, создающих негативное контрастирование в-в капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов, иногда из полипептидов, гидрофильна, антигенна препятствует фагоцитозу бак. Многие бак образуют микрокапсулу — слиз образование толщиной менее 0,2 мкм, выявл лишь при электронной микроскопии. Капсула предохраняет бак от повреждений, высыхания, т к, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных ф-ров макроорг и бактериофагов.
Жгутики бак опред подвижность бактериал кл. Жгутики предст собой тонкие нити. Они состоят из 3х частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными. Дисками жгутики прикреплены к цитоплазматической мембране и клет стенке. При этом создается эффект электромотора со стержнем — ротором, вращающим жгутик. Жгутики состоят из белка — флагеллина. Число жгутиков у бак разных видов варьирует от 1го (монотрих), по всему телу (перитрих), лофотрихи имеют пучок жгутиков на 1ом из концов кл, Амфитрихи по 1му жгутику или пучку жгутиков на противоположных концах кл. Жгутики выявл с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки спец методами, основанными на протравливании и адсорбции различных в-в, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) — нитевидные образования, более тонкие и короткие, чем жгутики. Пили отходят от поверхности кл и состоят из белка пилина. Известно несколько типов пилей.
Пили общего типа: прикрепления к субстрату, питание и водно-солевой обмен. Половые пили создают контакт между кл, осущ между ними передачу генетической информации путем конъюгации.
пили IV типа: концы обладают гидрофобностью, в результате чего они закручиваются. Они обладают антигенными св-вами, осущ контакт бак с кл хозяином,. Многие пили явл рецепторами для бактериофагов. Обнаружить можно с помощью электронной микроскопии.
Споры — своеобразная форма покоящихся бак с грам+ типом строения клет стенки. Споры кислотоустойчивы, поэтому окрашиваются по методу по методу Циля—Нельсена в красный, а вегетативная кл — в синий цвет. Форма спор бывает овальной и шаровидной, расположение в кл — терминальное, т.е. на конце палочки, субтерминальное — ближе к концу палочки и центральное. Процесс спорообразования (споруляция) проходит ряд стадий, в течение кот часть цитоплазмы и хромосома бактериал вегетативной кл отделяются, окружаясь врастающей цитоплазматической мембраной, — образуется проспора. В протопласте проспоры находятся нуклеоид, белоксинтезирующая сис-ма и сис-ма получения энергии, основанная на гликолизе. Цитохромы отсутст даже у аэробов. Не содержится АТФ, энергия для прорастания сохр в форме 3-глицеринфосфата. Проспору окружают 2 цитоплазматические мембраны. Слой, окруж внутр мембрану споры, называется стенкой споры, он состоит из пептидогликана и явл главным ист клет стенки при прорастании споры. Между наруж мембраной и стенкой споры формируется толстый слой, состоящий из пептидогликана, имеющего много сшивок, — кортекс. Кнаружи от внеш цитоплазматической мембраны располож оболочка споры, состоящая из кератиноподобных белков, содержащих множественные внутримолекулярные дисульфидные связи. Эта оболочка обеспечивает резистентность к химич агентам. Споры некот бак имеют дополнит покров — экзоспориум липопротеиновой природы. Т о формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой к-ты и ионов кальция. Спора приобретает термоустойчивость, кот связывают с наличием в ней дипиколината кальция. Спора долго может сохр из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, (З-оксимасляной к-ты и полифосфатов (волютин). Они накапливаются при избытке питат в-в в окр среде и выполняют роль запас в-в для питания и энергетич потребностей. Волютин обладает сродством к основным красителям и легко выявл с помощью спец методов окраски (н, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в краснофиолетовый цвет, а цитоплазма бак — в синий. Характерное распол гранул волютина выявл у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов кл. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронноплотных гранул размером 0,1 — 1 мкм.
7. Спорообразование у бак. Биологическая роль спор, располож спор в бактериал кл. Как называются палочки, образующие споры; каким способом окрашивают споры. Назовите заб, возбуд кот явл спорообразующие бак.
Превращение бактериал кл в спору происходит при попадании бак во внеш среду, чаще - в почву.
Причины перехода бак к спорообразованию: недостаток питат в-в; недостаток воды; накопление продуктов обмена и др неблагоприятные условия.
В оптимал усл происходит проростание споры. Сначала она активно поглощает воду и набухает, усил дыхание, возрастает активность ферментов. Затем спора лопается и из неё выходит вегетативная кл.
Биологическая роль спор - Сохр наследств инф-ции при неблагоприятных условиях внешней среды. Споры у бактерий не служат для размножения!
Среди бак споры образуют: бациллы (спора не превышает диаметр вегетативной кл), клостридии (спора превышает диаметр вегетативной кл).
Распол спор в кл бак: центрально (в центре), субтерминально (ближе к концу), терминально (на конце кл).
Споры кислотоустойчивы. Для их окраски испол метод Ожешко (Ауески) или Циля-Нильсена. При этом эндоспоры окрашиваются в красный цвет, а вегетативные кл - в сине-фиолетовый цвет
Ботулизм, столбняк, газовая гангрена, сибирская язва и т.д.