170. Синтез белка на рибосомах. Условия необходимые для реализации этого процесса.
171. Трансляция - как процесс реализации генетической информации в структурах синтезируемых на рибосомах полипептидныx цепей. Это биосинтез белка на матрице мРНК. Синтез начитнается после переноса инфо и ДНК на мРНК, которая несет инфо об одной полипептидной цепи. Есть 3 основных стадии трансляции: инициация, элонгация, терминация. Во время начальной стадии биосинтеза белков, инициации, обычно метиониновый кодон узнаётся малой субъединицей рибосомы, к которой при помощи белковых факторов инициации присоединена метиониновая транспортная РНК (тРНК). После узнавания стартового кодона к малой субъединице присоединяется большая субъединица и начинается вторая стадия трансляции — элонгация. При каждом движении рибосомы от 5' к 3' концу мРНК считывается один кодон путём образования водородных связей между тремя нуклеотидами (кодоном) мРНК и комплементарным ему антикодоном транспортной РНК, к которой присоединена соответствующая аминокислота. Синтез пептидной связи катализируется рибосомальной РНК (рРНК), образующей пептидилтрансферазный центр рибосомы. Рибосомальная РНК катализирует образование пептидной связи между последней аминокислотой растущего пептида и аминокислотой, присоединённой к тРНК, позиционируя атомы азота и углерода в положении, благоприятном для прохождения реакции. Ферменты аминоацил-тРНК-синтетазы присоединяют аминокислоты к их тРНК. Третья и последняя стадия трансляции, терминация, происходит при достижении рибосомой стоп-кодона, после чего белковые факторы терминации гидролизуют последнюю тРНК от белка, прекращая его синтез. Таким образом, в рибосомах белки всегда синтезируются от N- к C-концу.
172. Распад пуриновых оснований. Химизм процесса и его медико-биологическое значение. Подагра.
Образование мочевой к-ты происх.в печени и в кишечнике. У чел-ка моч.к-та явл.конечным продуктом обмена пуринов и выводится с мочей. В норме содержание моч.к-ты в сут.моче-400-600мг/дл. А в крови – 3-7-мг/дл. Увеличение-гиперурикимия. Подагра – заболевание, при котором мочевая к-та и ураты из-за плохой растворимости начин.кристализоваться и откладываться в суставных хрящах, связказ и мяг.тканях, вызывая воспаление суставов и мочекаменную болезнь. Генетически болезнь вызвана дефектами -ФРДФ-снтетазы, связанными с гиперактивацией фермента/ -частичной потерей активности гипоксантин-гуанинфосфорибозилтрансферазы, обеспечивая повторное исп.пуринов.
173. Распад пуриновых оснований. Химизм процесса и его медико-биологическое значение. Подагра.(см.172)
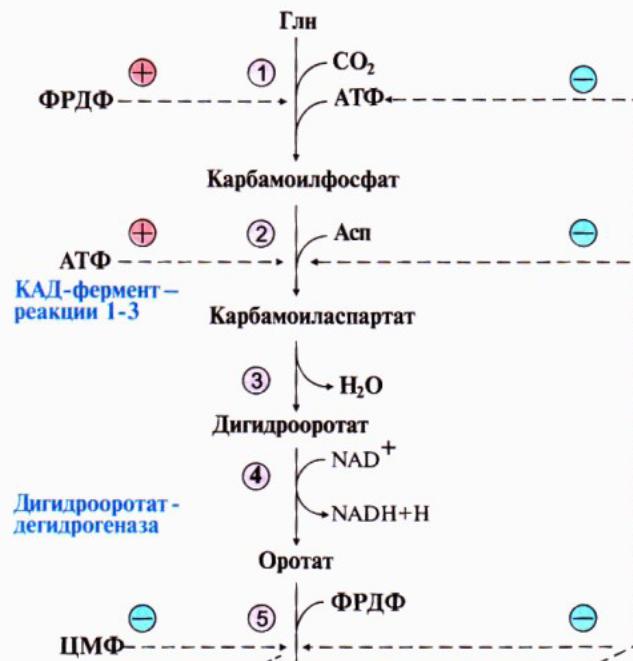
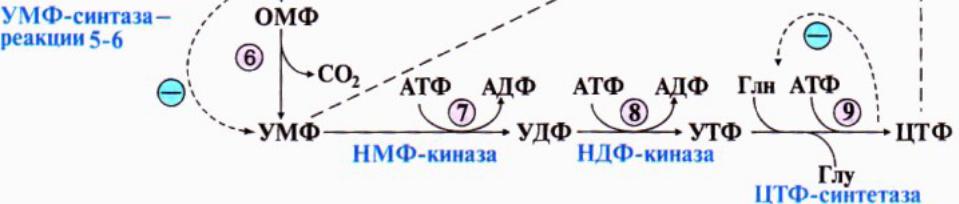
174. Представления о распаде и биосинтезе пиримидиновых нуклеотидов. УМФ явл.общим предшественником ост.пиримидиновых оснований. Синтез УМФ de novo вкл.шесть реакций, протекает в цитозоле клеток при участии 3хферментов. Пиримидиновое кольцо собирается из глутамина, аспартата и СО2, и после окисления превращается в оротат.из оротата в нуклеотид идет под действием ФРДФ.
Регуляция процесса: аллостерически по механизму отрицат.обратной связи конечными продуктами УМФ и ЦТФ. Синтез пуриновых и пиримидиновых оснований строго координируется: ФРДФ активирует оба синтеза, а накопление нуклеотидов ингибирует образ.ФРДФ отриц.обратной связью.
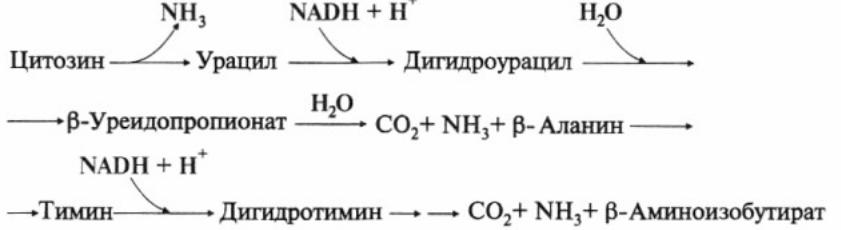
Катаболизм: