

12. Вітамін н (біотин): біологічні властивості, механізм дії
антисеборейний
Суточная потребность 150-200 мкг.
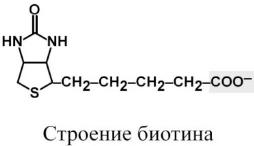
Строение
Гетероциклическая часть молекулы состоит из имидазольного и тиофенонового циклов. К последнему присоединена валериановая кислота, которая через COOH-группу связывается с лизином белковой части молекулы. Биотин-лизиновый конъюгат носит название биоцитин.
Биохимические функции
Биотин участвует в переносе СО2 либо из НСО3– (реакции карбоксилирования), либо от R-СООН (реакция транскарбоксилирования). Этот перенос необходим:
при синтезе оксалоацетата – биотин находится в составе пируваткарбоксилазы, что обеспечивает поддержание активности цикла трикарбоновых кислот и глюконеогенеза,
в синтезе жирных кислот – биотин находится в составе ацетил-SКоА-карбоксилазы,
в утилизации разветвленных углеродных цепей при катаболизме лейцина, изолейцина и некоторых жирных кислот – находится в составе пропионил-SКоА-карбоксилазы, образующей метилмалонил-SКоА. В дальнейшем метилмалонил-SКоА метаболизирует в реакции изомеризации с участием витамина B12.
Гіповітаміноз. У людей недостатність частіше пов’язана з порушенням функціонування мікрофлори кишечнику (застосування антибіотиків та сульфаніламіді, довге парентеральне харчування). Проявами гіповітамінозу є дерматити себорейного типу, може спостерігатись випадіння волосся, ураження нігтів, м’язова слабкість, анемія.
Антивитамины: гликопротеин авидин (антивитамин, связывающий биотин).
13. Вітамін в3 (пантотенова кислота): біологічні властивості, механізм дії
Антидерматитний
Суточная потребность 10-15 мг.
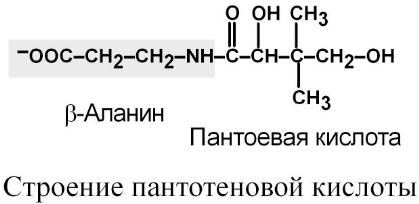
Строение
Витамин существует только в виде пантотеновой кислоты, в ее составе находится β-аланин и пантоевая кислота (2,4-дигидрокси-3,3-диметилмасляная).
Его коферментными формами являются кофермент А (коэнзим А, HS-КоА) и 4-фосфопантетеин.
Вітамін В3 виконує лише коферментні функції.
1. Фосфопантетеїн є активною субодиницею АПБ (ацил-переносного білка), що входить до складу синтази жирних кислот.
2. КоА-SH як кофермент ІІ групи забезпечує:
Активацію ацетату з утворенням ацетил-КоА, який необхідний для синтезу жирних кислот, кетонових тіл, холестерину, стероїдних гормонів, ацетилхоліну та ацетилглюкозамінів.
·Активацію жирних кислот з утворенням ацил-КоА ефірів.
·Окисне декарбоксилювання α-кетокислот (піровиноградної та α–кетоглутарової).
·Забезпечує реакції знешкодження ксенобіотиків.
Гіповітаміноз.
При недостатністі виникають дерматити (випадіння волос, зупинка росту, ураження слизової оболонки, неврити, ураження наднирників). Вторинна недостатність вітаміну розвивається при багатьох хронічних захворюваннях, тривалому застосуванні діуретиків і алкоголізмі.
Клиническая картина. Проявляется в виде педиолалгии (эритромелалгии) – поражение малых артерий дистальных отделов нижних конечностей, симптомом является жжение в стопах. В эксперименте проявляется поседение волос, поражение кожи и ЖКТ, дисфункции нервной системы, дистрофия надпочечников, стеатоз печени, апатия, депрессия, мышечная слабость, судороги. Но так как витамин есть во всех продуктах, гиповитаминоз встречается очень редко.
Антивитамины пантотеновой кислоты (B5). Одним из самых сильных антивитаминов является α-метилпантотеновая кислота. Она вызывает выраженные признаки недостаточности витамина в виде периферических нефропатий и нарушений функции коры надпочечников.