

31. Ферменти біологічного окислення в мітохондріях: піридин-, флавін-залежні дегідрогенази, цитохроми
1. Дегідрогенази, залежні від нікотинамідних коферментів (НАД(Ф)-залежні дегідрогенази).
Коферментами цих дегідрогеназ є нуклеотиди НАД або НАДФ, у структурі молекул яких міститься похідна піридину - нікотинамід. Зв'язок між НАД (або НАДФ) та білковою частиною ферменту (апоферментом) у складі піридинзалежних дегідрогеназ нестійкий: він утворюється та руйнується в процесі каталітичного циклу, що дозволяє вважати нікотинамідні нуклеотиди скоріше субстратами, ніж простетичними групами.
А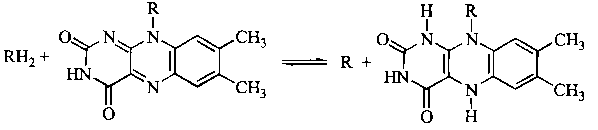 ктивною
структурою в молекулі НАД або НАДФ, що
акцептує відновлювальні еквіваленти
від субстрату, є піридинове кільце
никотинаміду. У ході ферментативної
реакції субстрат відщеплює два атоми
водню (2Н+
+ 2е-),
один з яких у формі гідридіона: Н-
(тобто Н+
+ 2е-)
приєднується до піридинового кільця
НАД(Ф), а другий у вигляді протона (іона
Н+)
надходить у реакційне середовище. Як
свідчить наведене рівняння, під час
реакції до четвертого вуглецевого атома
нікотинаміду приєднується атом водню
(тобто Н++
е-),
а додатковий електрон гідридіона
взаємодіє з азотом піридинового кільця.
Дегідрогенази, залежні від нікотинамідних
коферментів, дуже поширені в живих
клітинах. Вони виконують функції
анаеробних дегідрогеназ, що відщеплюють
протони та електрони від багатьох
субстратів, відновлюючи НАД або НАДФ,
які передають в подальшому відновлювальні
еквіваленти на інші акцептори. НАД-залежні
дегідрогенази - ці ферменти каталізують
окислювально-відновлювальні реакції,
що містяться на окислювальних шляхах
метаболізму - гліколізу, циклу лимонної
кислоти, β-окислення жирних кислот,
окисного дезамінування амінокислот,
дихального ланцюга мітохондрій.
НАДФ-залежні дегідрогенази - ці ферменти
беруть участь у процесах відновлювального
синтезу, що відбуваються в цитозолі,
зокрема постачають атоми водню при
синтезі жирних кислот та стероїдів.
Головним джерелом відновленого НАДФ є
дегідрогеназні реакції пентозофосфатного
шляху окислення глюкози.
ктивною
структурою в молекулі НАД або НАДФ, що
акцептує відновлювальні еквіваленти
від субстрату, є піридинове кільце
никотинаміду. У ході ферментативної
реакції субстрат відщеплює два атоми
водню (2Н+
+ 2е-),
один з яких у формі гідридіона: Н-
(тобто Н+
+ 2е-)
приєднується до піридинового кільця
НАД(Ф), а другий у вигляді протона (іона
Н+)
надходить у реакційне середовище. Як
свідчить наведене рівняння, під час
реакції до четвертого вуглецевого атома
нікотинаміду приєднується атом водню
(тобто Н++
е-),
а додатковий електрон гідридіона
взаємодіє з азотом піридинового кільця.
Дегідрогенази, залежні від нікотинамідних
коферментів, дуже поширені в живих
клітинах. Вони виконують функції
анаеробних дегідрогеназ, що відщеплюють
протони та електрони від багатьох
субстратів, відновлюючи НАД або НАДФ,
які передають в подальшому відновлювальні
еквіваленти на інші акцептори. НАД-залежні
дегідрогенази - ці ферменти каталізують
окислювально-відновлювальні реакції,
що містяться на окислювальних шляхах
метаболізму - гліколізу, циклу лимонної
кислоти, β-окислення жирних кислот,
окисного дезамінування амінокислот,
дихального ланцюга мітохондрій.
НАДФ-залежні дегідрогенази - ці ферменти
беруть участь у процесах відновлювального
синтезу, що відбуваються в цитозолі,
зокрема постачають атоми водню при
синтезі жирних кислот та стероїдів.
Головним джерелом відновленого НАДФ є
дегідрогеназні реакції пентозофосфатного
шляху окислення глюкози.
2. Флавінзалежні дегідрогенази
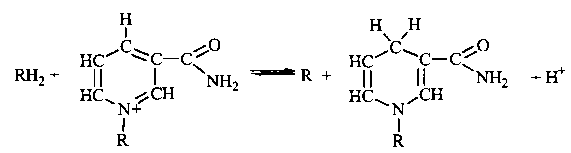 Дегідрогенази
цього типу за хімічною природою є
флавопротеїнами, простетичними групами
в яких є флавінаденіндинуклеотид (ФАД)
та флавінмононуклеотид (ФАД). На відміну
від піридинзалежних дегідрогеназ, у
більшості флавінзалежних ферментів
коферменти (ФАД та ФМН) міцно зв'язані
з білковою частиною і не відщеплюються
від неї на жодній стадії каталітичного
циклу. Виключенням є ФАД-залежна оксидаза
D-амінокислот, у складі якої білок має
низьку спорідненість із коферментом.
Активною частиною молекули ФАД або ФМН,
що бере участь в окислювально-відновлювальній
реакції, є ізоалоксазинове кільце
рибофлавіну, яке акцептує два атоми
водню (2Н+
+ 2е-)
від субстрату.
Дегідрогенази
цього типу за хімічною природою є
флавопротеїнами, простетичними групами
в яких є флавінаденіндинуклеотид (ФАД)
та флавінмононуклеотид (ФАД). На відміну
від піридинзалежних дегідрогеназ, у
більшості флавінзалежних ферментів
коферменти (ФАД та ФМН) міцно зв'язані
з білковою частиною і не відщеплюються
від неї на жодній стадії каталітичного
циклу. Виключенням є ФАД-залежна оксидаза
D-амінокислот, у складі якої білок має
низьку спорідненість із коферментом.
Активною частиною молекули ФАД або ФМН,
що бере участь в окислювально-відновлювальній
реакції, є ізоалоксазинове кільце
рибофлавіну, яке акцептує два атоми
водню (2Н+
+ 2е-)
від субстрату.
Цитохроми
- залізовмісні білки мітохондрій, що
належать до класу гемопротеїнів. У
цитохромах іон заліза входить до складу
металопорфіринового комплексу (гемінове
залізо), близького за хімічною структурою
до простетичних груп гемоглобіну та
міоглобіну. За рахунок оберненої зміни
валентності гемінового заліза цитохроми
виконують функцію транспорту електронів
у ланцюгах біологічного окислення в
аеробних клітинах:
![]()
Залежно від характерних особливостей спектрів поглинання, розрізняють три класи цитохромів (а, b, с). У мітохондріях еукаріотів наявні п'ять різновидів цитохромів - b, с, с1, а, а3; в ендоплазматичному ретикулумі гепатоцитів містяться цитохроми Р-450 та b5 що беруть участь у реакціях окислювального гідроксилування.
бх до 40