

68. Трансамінування амінокислот: реакції та їх біохімічне значення, механізми дії амінотрансфераз.
Реакції трансамінування полягають у переносі а-аміногрупи від амінокислоти на а-вуглецевий атом а-кетокислоти - акцептора аміногрупи (здебільшого- а-кетоглутарату та пірувату); в результаті реакції утворюється а-кетоаналог вихідної амінокислоти та нова амінокислота. Ферменти, що каталізують реакції трансамінування, - амінотрансферази (трансамінази).
Найбільш
поширеними є такі амінотрансферази:

Реакції трансамінування, що каталізуються амінотрансферазами, активно перебігають в багатьох органах, найактивніше - в печінці, скелетних м'язах, міокарді, головному мозку, нирках. Визначення активності аланінамінотрансферази (аланінової трансамінази - АлАТ) та аспартатамінотрансферази (аспарагінової трансамінази - АсАТ) широко застосовується в медичній практиці з метою діагностики пошкоджень внутрішніх органів. Внаслідок виходу цих ферментних білків через ушкоджені клітинні мембрани в кров при інфаркті міокарда спостерігається значне підвищення активності в сироватці крові АсАТ, при вірусних та токсичних пошкодженнях печінки - АлАТ.
Механізм дії амінотрансфераз
Амінотрансферази є складними білками-ферментами, простетичною групою в яких
є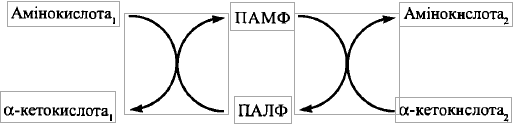 коферментні форми вітаміну В6 (піридоксину,
піридоксолу)- піридоксальфосфат (ПАЛФ)
та піридоксамінфосфат (ПАМФ), що
утворюється з ПАЛФ у процесі переносу
аміногрупи.
коферментні форми вітаміну В6 (піридоксину,
піридоксолу)- піридоксальфосфат (ПАЛФ)
та піридоксамінфосфат (ПАМФ), що
утворюється з ПАЛФ у процесі переносу
аміногрупи.
Процес складається з двох напівреакцій:
1) Взаємодія амінокислоти, що втрачає аміногрупу, з піридоксальфосфатом з утворенням кетокислоти та піридоксамінфосфату:
Механізм реакції полягає у взаємодії амінокислоти з ПАЛФ-Е із заміною альдимінного зв'язку у складі ПАЛФ-Е на альдимінний зв' язок між коферментом та амінокислотою; після внутрішньомолекулярного перегрупування відбувається гідроліз кетиміну з утворенням коферментної форми, що містить аміногрупу (ПАМФ), та а-кетокислоти
2) Взаємодія а-кетокислоти, що акцептує аміногрупу, з піридоксамінфосфатом з утворенням нової амінокислоти та регенерацією піридоксальфосфату:
Механізм цієї напівреакції аналогічний розглянутому для першої напівреакції (при її перебігу у зворотному напрямку); ПАЛФ, що регенерує в результаті другої напівреакції, знову сполучається альдимінним зв'язком з білковою частиною ферменту.
Біохімічне значення реакцій трансамінування - в реакціях трансамінування не відбувається дезамінування, тобто вивільнення аміаку, оскільки аміногрупа, що відщеплюється від а-L-амінокислоти, акцептується відповідною а-кетокислотою
У печінці роль трансамінування полягає в його колекторній функціі: тобто збиранні аміногруп від різних амінокислот переважно в одній молекулярній формі - у вигляді L-глутамінової кислоти. Біохімічний сенс такого процесу полягає в тому, що саме L-глутамат є основним субстратом реакцій дезамінування, тобто постачальником аміногруп на метаболічний шлях утворення сечовини - кінцевого продукту азотистого катаболізму.
У м'язах спрямованість реакцій трансамінування призводить до утворення значної кількості аланіну (за рахунок переамінування амінокислот з піруватом), що виділяється у кров'яне русло і поглинається гепатоцитами; в печінці аланін знову перетворюється на піруват, який використовується в глюконеогенезі (глюкозо-аланіновий цикл)