

1. Піруват у фосфоенолпіруват. – у дві стадії,
1) піруват+СО2+АТФ-> оксалоацетат+ АДФ + Фн. Ферм. піруваткарбоксилаза
2) оксалоацетат + ГТФ -> фосфоенолпіруват+СО2+ГДФ. Ферм. ФЕП-карбоксикіназа (у цитозолі та мітохондріях)
Транспорт оксалоацетату з мітох. в цитозоль – через човникові системи: малатна(основна, малатдегідрогеназа), аспартатна(аспартатамінотрансфераза), цитратна(цитратсинтаза)
2. Фруктозо-1,6-дифосф. У фруктозо-6-фосф.:
фрукт.-1,6-дифосф. + Н2О -> фрукт.-6-фосфат + Фн. Ферм. фруктозо-1,6-дифосфатаза (у печінці, нирках, епітеліоцитах кишечника)
3. Глюкозо-6-фосф. У глюкозу:
глюк-6-фосф.+Н2О -> глюкоза + Фн. Ферм. глюкозо-6-фосфатаза (у мембранах ЕПС гепатоцитів).
Отже, для синтезу 1 глюкози з 2 піруватів треба 6 макроерг. зв’язків
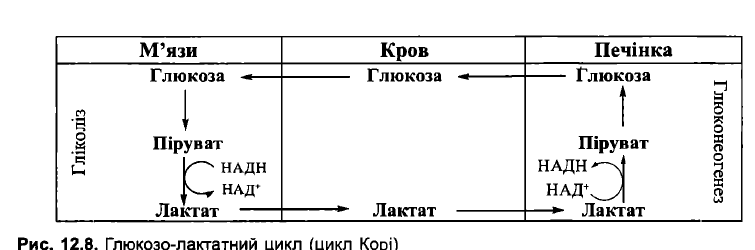
47. Глюкозо-лактатний (цикл Корі) та глюкозо-аланіновий цикли.
Глюкозо-лактатний (цикл Корі) пов’язує процеси утворення лактату в м’язовій тканині (анаер.гліколіз), його вихід у кров через плазмат.мембрани та використання лактату (після реокислення в піруват) у гепатоцитах для глюконеогенезу.
За
рахунок відновлення пірувату в лактат
у м’язах та його подальшого використання
в печінці, м’язи не тільки втрачають
«зайву» мол.кислот, яка утв. при
інтенс.фіз.праці, а й підтримують високий
НАД/НАДН (для активного гліколізу).
Глюкозо-аланіновий цикл:
А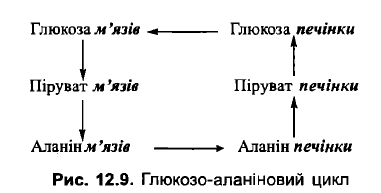 ланін,
який може утворюватися у скелетних
м'язах в оборотній реакції трансамінування
пірувату з глутаматом:
ланін,
який може утворюватися у скелетних
м'язах в оборотній реакції трансамінування
пірувату з глутаматом:
Піруват + Глутамат -> Aланін + а-кетоглутарат
Вивільняючись із працюючих м'язів у кров, аланін поглинається гепатоцитами і (після перетворення в піруват) використовується в глюконеогенезі (глюкозо-аланіновий цикл)
48. Глюкоза крові (глюкоземія): нормоглікемія, гіпо- та гіперглікемії, глюкозурія.
Цукровий діабет – патологія обміну глюкози.
нормоглікемія 3,3 - 5,5 ммоль\л
гіпоглікемія 2.5 и ниже
гіперглікемія више 6
Причини гіпоглікемії:
- Прийом інсуліну ( інсулін активує глікогенез)
- Первинні ензимопатії:
галактоземія ( галактокінази)
непереносимість фруктози (дефіцит фруктозо-1-фосфатальдолази)
- Глікогеноз (печінковий) ( глікоген не може розпадатися, глюкоза не утв, глікогенсинтаза)
- Інсулома (гіперінсулінізм)
- Наднирникова недостатність (хвороба Аддісона) гормони, що стимулюють глікогену
- Гіпофізарна недостатність
- Захворювання ШКТ (мала абсорбція моноз)
УСКЛАДНЕННЯ
Гіпоглікемічна кома – це втрата свідомості за рахунок критичного гіпоергозу нейронів.
Причини гіперглікемії:
• Аліментарна (фізіологічна)
• Стресорна
• Ураження ЦНС
• Печінкова недостатність (цироз)
( с глюкозы синтезируется гликоген,цирроз печени - накапливается глюкоза)
• Ендокринна:
Цукровий діабет
Іценко-Кушинга (гіперкортицизм)
Глюкагонома
Феохромацитома
Гіпофізарний гігантизм
Гіпертиреоз
УСКЛАДНЕННЯ
Діабетична гіперглікемічна кома
Гіперглікемічна гіперосмолярна кома.
Глюкозурі́я — наявність глюкози в сечі.
ГЛЮКОЗУРІЯ
У нормі сеча не містить глюкози, оскільки нирки здатні реабсорбувати (повертати до кровотоку) весь обсяг глюкози, що пройшов через нирковий клубочок в просвіт канальців нефрона. У переважній більшості випадків глюкозурія є симптомом декомпенсованого цукрового діабету як результат патологічного збільшення концентрації глюкози в крові. Рідкісним винятком є порушення реабсорбції в самій нирці — так звана ниркова глюкозурія. Глюкозурія веде до надмірної втрати води з сечею та дегідратації організму.
При перевищенні концентрації глюкози в крові вище ниркового порогу (8,88-9,99 ммоль / л) глюкоза починає надходити в сечу, і виникає глюкозурія
Цукр.Діаб – генетично обумовлений дефіцит інсуліну: зниження продукції бета-клітинами підшлункової залози або нездатність клітинних рецепторів реагувати на інсулін.
1)Інсулінозалежний – внаслідок руйнування (звичайно більше 90%) б-клітин через автоімунний процес. 10-15% усіх випадків, проявляється гіперглікемією та схильністю до кетонемії та кетоацидозу.
2)Інсулінонезалежний – збереження бета-клітин, але порушення специфічних реакцій на дію інсуліну або регуляцію його секреції під впливом підвищеної конц. глюкози крові.