

Bykov-_gistologia_obschaya
.pdfМикроворсинки - пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. Мнкроворсинки обеспечивают многократное увеличение площади поверхности клетки, на которой происходит расщепление и всасывание веществ. На апикальной поверхности некоторых клеток, активно участвующих в указанных процессах (в эпителии тонкой кишки и почечных канальцев) имеется до нескольких тысяч микроворсинок, образующих в совокупности щеточную каемку.
Рис 3-17. Схема ультраструкгурной организации микроворсинки. АМФ - актиновые микрофиламенты, АВ - аморфное вещество (апикальной части микроворсинки), Ф, В - фимбрин и виллин (белки, образующие поперечные сшивки в пучке АМФ), ММ - молекулы минимиозина (прикрепляющие пучок АМФ к плазмолемме микроворсинки), ТС - терминальная сеть АМФ, С - спектриновые мостики (прикрепляют ТС к плазмолемме), МФ - миозиновые филаменты, ПФ - промежуточные филаменты, ГК - гликокаликс.
Каркас каждой микроворсинки образован пучком, содержащим около 40 микрофиламентов, лежащих вдоль ее длинной оси (рис. 3-17). В апикальной части микроворсинки этот пучок закреплен в аморфном веществе. Его жесткость обусловлена поперечными сшивками из белков фимбрина и виллина, изнутри пучок прикреплен к плазмолемме микроворсинки особыми белковыми мостиками (молекулами минимиозина). У основания мнкроворсинки микрофиламенты пучка вплетается
- 71 -
в терминальную сеть, среди элементов которой имеются миозиновые филаменты. Взаимодействие актиновых и миозиновых филаментов терминальной сети, вероятно, обусловливает тонус и конфигурацию микроворсинки.
Стереоцилии - видоизмененные длинные (в некоторых клетках - ветвящиеся) микроворсинки - выявляются значительно реже, чем микроворсинки и, подобно последним, содержат пучок микрофиламентов.
Промежуточные филаменты
Промежуточные филаменты - прочные и устойчивые в химическом отношении белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микро-филаментов). Они встречаются в клетках разных тканей (см. ниже) и располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, входят в состав десмосом и полудесмосом эпителиальных клеток (в плазмолемме которых они закреплены посредством трансмембранных белков), лежат по всей длине отростков нейронов. Промежуточные филаменты образованы нитевидными белковыми молекулами, сплетенными друг с другом наподобие каната.
Функции промежуточных филаментов изучены недостаточно;
установлено, однако, что они не влияют ни на движение, ни на деление клетки.
Ких основным функциям относятся:
(1)структурная - поддерживающая и опорная, обеспечение распределения органелл по определенным участкам цитоплазмы;
(2)обеспечение равномерного распределения сил деформации между клетками ткани, что препятствует повреждению отдельных клеток (благодаря связи промежуточных филаментов с трансмембранньгми белками десмосом и полудесмосом);
(3)участие в образовании рогового вещества в эпителии кожи; в
эпителиальных клетках связываются с другими белками и образуют непроницаемые барьеры (роговые чешуйки), являются главным компонентом волос и ногтей;
(4)поддержание формы отростков нервных клеток и фиксация трансмембраниых белков (в частности, ионных каналов);
(5)удержание миофибрилл в мышечной ткани и прикрепление их к плазмолемме. что обеспечивает их сократительную функцию.
Очевидно, что функции, отмеченные цифрами (2)-(5), служат лишь частными проявлениями более общей структурной функции (1) в различных тканях.
-72 -
В поврежденной клетке сеть промежуточных филаментов (в отличие от других компонентов цитоскелета) спадается и концентрируется вокруг ядра, связывая поврежденные органеллы и белковые агрегаты. Формируется своеобразная структура, которая наподобие кокона концентрирует поврежденные компоненты клетки для последующего уничтожения путем их внутриклеточного переваривания. В ходе восстановления структуры и функции клетки после повреждения сеть промежуточных филаментов вновь развертывается по всей цитоплазме. В отличие от микрофиламентов и микротрубочек, для образования промежуточных филаментов не требуется АТФ, причем они не подвергаются постоянной сборке и диссоциации, а представляют собой менее лабильные и сравнительно устойчивые структуры.
Распределение промежуточных филаментов различных классов в клетках и тканях человека
Классы промежуточных |
Типы клеток и тканей |
филаментов |
|
|
|
(цито-)кератиновые |
эпителиальные |
(тонофиламенты) |
|
десминовые |
мышечные ткани - гладкие |
|
(кроме миоцитов сосудов) и |
|
поперечнополосатые |
виментиновые |
различные клетки мезенхимного |
|
происхождения: фибробласты, |
|
макрофаги, остеобласты, |
|
хондробласты, эндотелий и |
|
гладкие миоциты сосудов |
|
|
нейрофиламенты |
нейроны |
глиальные (содержат |
глиальные клетки (астроциты, |
глиальный |
олигодендроглиоциты) |
фибриллярный кислый |
|
ламины (образуют |
все типы клеток |
кариоскелет) |
|
- 73 -
Классы промежуточных филаментов и их идентификация.
Несмотря на то, что строение промежуточных филаментов в клетках различных типов сходно, они существенно различаются по своей молекулярной массе и химической природе, что может быть продемонстрировано иммуноцитохимическими методами с антителами к промежуточным филаментам различных классов. Различают 6 основных классов промежуточных филаментов (см. выше). В цитоплазме большинства клеток содержится лишь один их класс; в части клеток выявляются два класса, из которых один является основным.
Идентификация классов промежуточных филаментов имеет важное значение в диагностике опухолей для выявления тканевой принадлежности опухолевых клеток, что может определить выбор лечения и прогноз. Наибольшее диагностическое значение имеет выявление цитокератинов, десмина и глиального фибриллярного кислого белка, которые служат маркерами опухолей эпителиального, мышечного и глиального происхождения. Менее отчетливые результаты дает обнаружение виментина, который экспрессируется и коэкспрессируется (экспрессируется в сочетании с белками других классов промежуточных филаментов) многими типами клеток. Существенную информацию о степени поражения эпителия можно получить путем определения экспрессии молекулярных форм кератинов, специфичных для клеток конкретной локализации и уровня дифференцировки. Таким путем можно установить, например, ранние предраковые изменения в эпителии, не выявляемые стандартными морфологическими методами.
Микротрабекулы
Микротрабекулы - наименее изученная система цитоскелета, само существование которой оспаривается многими исследователями. Предполагают, что гри описанные выше системы филаментов пронизываются и объединяются некоей четвертой системой, названной микротрабекулярной сетью. Последняя выявляется при высоковольтной электронной микроскопии как система нитей неравномерной толщины (2-10 нм), связывающая три системы цитоскелета, различные органеллы и плазмолемму. В "узлах" микротрабекулярной сети располагаются свободные рибосомы и полисомы. Белок, образующий микротрабекулярную сеть, не идентифшшрован. Высказываются предположения о том, что эта сеть представляет собой артефакт, возникающий в результате преципитации и коагуляции белков при фиксации цитоплазмы клетки.
- 74 -
ВКЛЮЧЕНИЯ
Включения цитоплазмы - временные ее компонента, обусловленные накоплением продуктов метаболизма клеток. Традиционно подразделяются на трофические, секреторные, экскреторные и пигментные.
Трофические включения разделяют в зависимости от природы накапливаемого вещества. Липидные включения встречаются в виде липидных капель (особенно крупных в жировых клетках), которые располагаются в цитоплазме по отдельности или сливаются друг с другом. Их вид на электронно-микроскопических фотографиях варьирует в зависимости от способа фиксации. На гистологических препаратах они обычно имеют вид светлых ("пустых") вакуолей, так как при стандартных методах обработки ткани липиды растворяются. Липидные капли служат источником веществ, используемых в качестве энергетических субстратов; в некоторых клетках (например, продуцирующих стероидные гормоны) они могут содержать субстраты, необходимые для последующего синтеза. Из углеводных трофических включений наиболее распространены гранулы гликогена, представляющего собой полимер глюкозы. Они встречаются в виде плотных гранул диаметром 20-30 нм (β-частиц), которые часто образуют скопления (розетки), называемые α-частицами (см. рис. 3-13). Гранулы гликогена часто расположены вблизи аЭПС и используются в качестве источника энергии.
Секреторные включения обычно имеют вид мембранных пузырьков, содержащих секретируемый клеткой продукт; в мембране могут находиться ферменты, осуществляющие конечный процессинг продукта по мере перемещения пузырька к плазмолемме. Избыток невостребованного секреторного продукта поглощается и разрушается в цитоплазме клетки механизмом кринофагии (см. выше).
Экскреторные включения по своему строению сходны секреторными, однако они содержат вредные продукты метаболизма, подлежащие удалению из клетки.
Пигментные включения представляют собой скопления эндогенных или экзогенных пигментов, которые могут окружаться мембраной. К наиболее распространенным эндогенным пигментам относятся гемоглобин (растворен в цитоплазме эритроцитов, переносит кислород), гемосидерин (продукт обмена гемоглобина, накапливается в макрофагах в виде мелких плотных частиц ферритина), меланин (синтезируется в пигментных клетках - меланоцитах, в которых он накапливается и хи-
- 75 -
мически дозревает в окруженных мембраной гранулах - меланосомах), липофусцин (пигмент старения, накапливается в виде мембранных гранул с плотным содержимым, в котором определяются липидные капли).
ЯДРО КЛЕТКИ
Ядро является важнейшим компонентом клетки, содержащим ее генетический аппарат.
Функции ядра:
1)хранение генетической информации (в молекулах ДНК, находящихся в хромосомах);
2)реализацию генетической информации, контролирующей осуществление разнообразных процессов в клетке - от синтетических до запрограммированной гибели (аноптоза);
3)воспроизведение и передачу генетической информации (при делении
клетки).
Обычно в клетке имеется только одно ядро, однако встречаются многоядерные клетки, которые образуются вследствие деления клеток, не сопровождающегося цитотомией, или слияния нескольких одно-ядерных клеток (последние правильнее называть симпластами).
Форма ядра различных клеток неодинакова: встречаются клетки с округлым, овальным, бобовидным, палочковидным, многолопастным, сегментированным ядром; нередко на поверхности ядра имеются вдавления. Чаще всего форма ядра в целом соответствует форме клетки: оно обычно сферическое в клетках округлой или кубической формы, вытянутое или эллипсоидное в призматических клетках, уплощенное - в плоских.
Расположение ядра варьирует в разных клетках; оно может лежать в центре клетки (в клетках округлой, плоской, кубической или вытянутой формы), у ее базального полюса (в клетках призматической формы) или на периферии (например, в жировых клетках).
Величина ядра относительно постоянна для каждого типа клеток, однако она может меняться в определенных пределах, увеличиваясь при усилении функциональной активности клетки и уменьшаясь при ее угнетении.
Компоненты ядра. В ядре неделящейся (интерфазной) клетки выявляются кариолемма (ядерная оболочка), хроматин, ядрышко и кариоплазма (ядерный сок). Как будет видно из дальнейшего изложения,
- 76 -
хроматин и ядрышко представляют собой не самостоятельные компоненты ядра, а являются морфологическим отражением хромосом, присутствующих в интерфазном ядре, но не выявляемых в качестве отдельных образований.
Ядерная оболочка
Ядерная оболочка (кариолемма) на светооптическом уровне практически не определяется; под электронным микроскопом обнаруживается, что она состоит из двух мембран - наружной и внутренней, - разделенных полостью шириной 15-40 нм (перинуклеарным пространством) и смыкающихся в области ядерных пор (рис. 3-18 и 3-19).
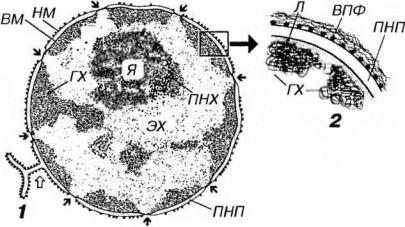
Рис. 3-18. Ядро клетки - общий вид (I) и участок ядерной оболочки (2). ГХ - гетерохроматин, ЭХ - эухроматин, Я - ядрышко, ПНХ - перинукпеолярный хроматин, НМ - наружная мембрана ядерной оболочки, ВМ - внутренняя мембрана, ПНП - перинуклеарное пространство, ВПФ - виментиновые промежуточные филаменты, Л - ламина. Черными стрелками показаны ядерные поры, белой - участок соединения ядерной оболочки с грЭПС.
Наружная мембрана составляет единое целое с мембранами грЭПС - на ее поверхности имеются рибосомы, а перинуклеарное пространство соответствует полости цистерн грЭПС и может содержать синтезированный материал. Со стороны цитоплазмы наружная мембрана окружена рыхлой сетью промежуточных (виментиновых) филаментов (см. рис. 3-18).
- 77 -
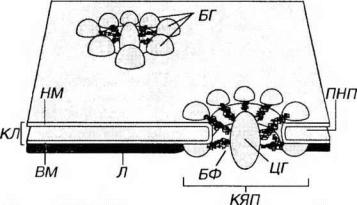
Рис. 3-19. Комплекс ядерной поры (КЯП). БГ - белковые гранулы, БФ - белковые фибриллы, ЦГ центральная гранула, КЛ - кариолемма: НМ - наружная мембрана (рибосомы на ее поверхности не показаны), ВМ - внутренняя мембрана кариолеммы, Л - ламина, ПНП - перинуклеарное пространство.
Внутренняя мембрана - гладкая, ее интегральные белки связаны с ядерной пластинкой - ламиной - слоем толщиной 80-300 нм, состоящим из переплетенных промежуточных филаментов (ламинов), образующих кариоскелет. Ламина играет очень важную роль в: (1) поддержании формы ядра;
(2) упорядоченной укладке хроматина; (3) структурной организации поровых комплексов; (4) формировании кариолеммы при делении клеток.
Ядерные поры занимают 3-35% поверхности ядерной оболочки. Они более многочисленны в ядрах интенсивно функционирующих клеток и отсутствуют в ядрах спермиев. Поры (см. рис. 3-19) содержат два параллельных кольца (по одному с каждой поверхности кариолеммы) диаметром 80 нм, которые образованы 8 белковыми гранулами. От этих гранул к центру сходятся фибриллы, формируйте перегородку (диафрагму) толщиной около 5 нм, в середине которой лежит центральная гранула (по некоторым представлениям, это - транспортируемая через пору субъединица рибосомы). Совокупность структур, связанных с ядерой порой, называется комплексом ядерной поры. Последний образует водный канал диаметром 9 нм, по которому движутся мелкие водорастворимые молекулы и ионы. Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации.
Ядерная оболочка в клетках животных и человека содержит до 2000-4000 поровых комплексов. В ядро из цитоплазмы через них поступают синтезированные белки, в обратном направлении переносятся молекулы РНК и субъединицы рибосом.
- 78 -
Функции комплекса ядерной поры:
1.Обеспечение регуляции избирательного транспорта веществ между цитоплазмой и ядром.
2.Активный перенос в ядро белков, имеющих особую маркировку в виде так называемой последовательности ядерной локализации - Nuclear Localization Sequence (NLS), распознаваемой рецепторами NLS (в комплексе поры).
3.Перенос в цитоплазму субъединиц рибосом, которые, однако, слишком велики для свободного прохождения пор; их транспорт, веро¬ятно, сопровождается изменением конформации порового комплекса.
Хроматин
Хроматин (от греч. chroma - краска) мелкие зернышки и глыбки материала, который обнаруживается в ядре клеток и окрашивается основными красителями. Хроматин состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Различают два вида хроматина - эухроматин и гетерохроматин.
Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции. Эти сегменты не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид гранул.
Таким образом, по морфологическим признакам ядра (соотношению содержания эу- и гетерохроматина) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки. При ее повышении это соотношение изменяется в пользу эухроматина, при снижении - нарастает содержание гетерохроматина. При полном подавлении функции ядра (например, в поврежденных и гибнущих клетках, при ороговении эпителиальных клеток эпидермиса - кератиноцитов, при образовании ретикулоцитов крови) оно уменьшается в размерах, содержит только гетерохроматин и окрашивается основными красителями интенсивно и равномерно. Такое явление называется кариопикнозом (от греч. karyon - ядро и pyknosis - уплотнение).
Распределение гетерохроматина (топография его частиц в ядре) и
соотношение содержания эу- и гетерохроматина характерны для клеток каждого типа, что позволяет осуществлять их идентификацию
- 79 -
как визуально, так и с помощью автоматических анализаторов изображения. Вместе с тем, имеются определенные общие закономерности распределения гетерохроматина в ядре: его скопления располагаются под кариолеммой, прерываясь в области пор (что обусловлено его связью с ламиной) и вокруг ядрышка (перинуклеолярный гетерохроматин), более мелкие глыбки разбросаны по всему ядру (см. рис. 3-18).
Тельце Барра - скопление гетерохрома гина, соответствующее одной Х- хромосоме у особей женского пола, которая в интерфазе плотно скручена и неактивна. В большинстве клеток оно лежит у кариолеммы, а в гранулоцитах крови имеет вид маленькой добавочной дольки ядра ("барабанной папочки"). Выявление тельца Барра (обычно в эпителиальных клетках слизистой оболочки полости рта) используется как диагностический тест для определения генетического пола (обязателен, в частности, для женщин, участвующих в Олимпийских Играх).
Упаковка хроматина в ядре. В деконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей каждую хромосому, равна в среднем, около 5 см, а общая длина молекул ДНК всех хромосом в ядре (диаметром около 10 мкм) составляет более 2 м (что сравнимо с укладкой нити длиной 20 км в теннисный мячик диаметром около 10 см), а в Б-период интерфазы - более 4 м. Конкретные механизмы, препятствующие спутыванию этих нитей во время транскрипции и репликации, остаются нераскрытыми, однако очевидна необходимость компактной упаковки молекул ДНК. В клеточном ядре это осуществляется благодаря их связи со специальными основными (гистоновыми) белками. Компактная упаковка ДНК в ядре обеспечивает:
(1)упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра;
(2)функциональный контроль активности генов (вследствие вли¬яния характера упаковки на активность отдельных участков генома.
Уровни упаковки хроматина (рис. 3-20). Начальный уровень упаковки хроматина, обеспечивающий образование нуклеосомной нити диаметром 11 нм, обусловлен намоткой двойной нити ДНК (диаметром 1 нм) на блоки дисковидной формы из 8 гистоновых молекул (нуклеосомы). Нуклеосомы разделены короткими участками свободной ДНК. Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити с формированием хроматиновой фибриллы диаметром 30 нм. В интерфазе хромосомы образованы хроматиновыми фибриллами, причем каждая хроматида состоит из одной фибриллы. При дальнейшей упаковке хроматиновые фибриллы образуют петли (петельные домены) диаметром 300 нм, каждый из которых соответствует одному или нескольким генам, а те, в свою очередь, в ре-
- 80 -