

Bykov-_gistologia_obschaya
.pdfСоотношение содержания в крови Т-лимфоцитов с маркерами CD4+ и
CD8+ оценивает общую активность иммунных реакций. При ВИЧ-инфекции, например, оно является важным диагностическим и прогностическим показателем. Снижение величины этого показателя с 2-3 (в норме) до 1 указывает на опасность иммуннодефицита (с вероятным развитием инфекций), а его падение ниже 0.5 обычно соответствует развернутой клинической картине СПИДа, осложненной тяжелыми инфекциями.
Т-киллеры (Тк), или Т-цитотоксические лимфоциты (ЦТЛ)
посредством ТКР и молекулы CD8 распознают эпитопы антигенов в комбинации с молекулами МНС I класса (рис. 8-4). Этот комплекс распознается Тк на АПК или на клетках-мишенях (зараженных вирусом или опухолевых). Тк живут меньше, чем Тх.
Рис. 8-4. Распознавание антигенов и активация Тк. Эпитоп антигена (АГ) в комплексе с молекулами МНС I класса (МНС I). который находится на поверхности клетки-мишени (КЛМ), распознается посредством ТКР и СD8 на мембране Тк, обеспечивая главный активационный сигнал (светлая звездочка). Дополнительная активация Тк достигается костимулирующим сигналом (черная звездочка) в результате взаимодействия костимулирующих молекул (КМ) на поверхности КЛМ с адгезионными молекулами (АМ) на поверхности Тк. При активации Тк выделяет ряд цитокинов.
Распознавание антигенов и активация Тк происходит под влиянием двух основных сигналов: (1) взаимодействия ТКР-СD8 на Тк с комплексом МНС I класса/эпитоп антигена на АПК или клетке-мишени; (2) воздействия цитокинов, выделяемых Тх, макрофагами и дендритными АПК.
- 231 -
Активированные Тк продуцируют ИФНγ и ИЛ-2, стимулируют представление антигенов, активируют клетки, участвующие в воспалении и, уничтожают клетки-мишени (зараженные вирусами, патогенными грибами и некоторыми бактериями), очищая от них организм. Их деятельность лежит также в основе противоопухолевого и трансплантационного иммунитета. Вещества, необходимые для уничтожения клеток-мишеней (перфорин, гранзимы и др.), накапливаются в образующихся 30-50 крупных (диаметром 0.5-2.0 мкм) цитоплазматических гранулах, покрытых мембраной и заполненных материалом, неоднородным по электронной плотности. По своему содержимому и морфологическим признакам эти гранулы сходны с гранулами NК-клеток.
При встрече с различными плетками Тк обследуют их поверхность в поисках антигенного эпитопа, который они способны распознать (в комплексе с молекулами МНС I класса). При обнаружении клетки-мишени зрелый Тк связывается с ней и оказывает на нее летальное цитотоксическое воздействие. После этого воздействия ("смертельного удара", или "смертельного поцелуя"), осуществляемого строго прицельно и не повреждающего соседние клетки. Тк отсоединяется от гибнущей клетки-мишени и ищет следующую жертву. Активность Тк находится под совместным контролем Тх и Тс; предполагают, что Тх действуют на Тк непосредственно, а Тс - косвенно (угнетая Тх).
Механизмы контактного цитотоксического действия Тк
("смертельного удара”), вызывающего гибель клетки-мишени, разрушают ее снаружи и изнутри (рис. 8-5):
1.Образование пор в плазмолемме клеток-мишеней. Тк вступает в контакт
склеткой-мишенью и в присутствии ионов Са2+ направленно (векторно) секретирует в межклеточное пространство особые белки перфорины, накопленные ранее в его цитоплазматических гранулах. Мономеры перфоринов встраиваются в качестве трансмембранных белков в плазмолемму клетокмишеней, а в дальнейшем образуют в ней агрегата в виде трансмембранных пор (рис. 8-6). Вследствие постоянного включения новых мономеров в агрегаты размеры пор растут, достигая 5-20 нм (внутренний диаметр наиболее крупных равен 10 нм). Поры, возникающие в плазмолемме при атаке Тк, очень сходны с образующимися при воздействии мембраyоатакуюшего комплекса компонентов комплемента. Формирование пор в плазмолемме приводит к нарушению осмотического равновесия клетки-мишени, ее набуханию и гибели. Образующиеся поры служат также проводниками веществ, вызывающих разрушение клеток-мишеней изнутри (см. ниже).
-232 -
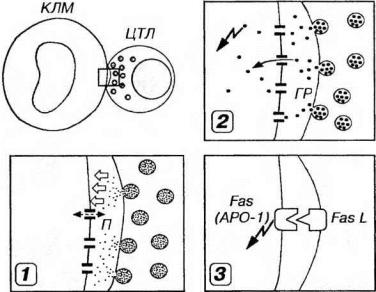
Рис. 8-5. Механизмы контактного действия цитотоксическогс лимфоцита (ЦТЛ) на клетку мишень (КЛМ). 1 - ЦТЛ секретирует в межклеточное пространство белки перфорины (П), которые встраиваются в плазмолемму КЛМ (белые стрелки) и образуют в ней трансмембранные поры, вызывающие нарушение осмотического равновесия КЛМ (пунктирная стрелка) и ее гибель (подробнее механизм образования пор показан на рис. 8-6). 2 - апоптоз КЛМ (стрелка в виде ’’молнии”) индуцируется в результате введения в ее цитоплазму через ранее образованные поры в плазмолемме ферментов гранзимое (ГР), синтезируемых и выделяемых ЦТЛ (стрелка) 3 - индук¬ия апоптоза КЛМ (стрелка в виде ’’молнии") в результате взаимодействия Fasлиганда (Fas-L) на поверхности Тк с антигеном Fas (АРО-1) на плазмолемме КЛМ. Указанные варианты взаимодействия не являются взаимоисключающими и осуществляются после того, как ЦТЛ посредством TKP и CD8 распознает комплекс антиген/ МНС I класса на поверхности КЛМ (механизм этого взаимодействия показан на рис. 8- 4).
2. Индукция апоптоза клеток-мишеней ферментами, введенными в их цитоплазму через поры в плазмолемме. Тк синтезируют и накапливают в гранулах ряд ферментов (из которых наиболее важна группа сериновых протеаз - гранзимов). Направленно выделяясь в межклеточное пространство при Са2+ - зависимом экзоцитозе гранул Тк, эти ферменты через образующиеся перфориновые поры проникают в цитоплазму клеток-мишеней и запускают программу апоптоза, вызывающую гибель этих клеток. При этом гранзимы действуют как прямо - на субстраты, расшепление которых непосредственно вызывает гибель клетки, так и косвенно, активируя цистеиновые протеазы семейства ICE (IL-1 Converting Enzyme - фермент, конвертирующий ИЛ-1), обусловливающие апоптоз.
- 233 -
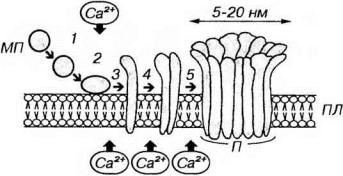
Рис. 8-6. Механизм образования пор в плазмолемме клетки-мишени при атаке Тк. 1 - мономеры перфорина (МП), выделяются Тх в пространство между ним и клеткой-мишенью; 2 - МП связываются с фосфолипидными головками плазмолеммы (ПЛ) клетки-мишени; 3 - МП претерпевают конформационные изменения и внедряются в липидный бислой; 4-5 - полимеризация МП с образованием поры (П) - агрегата МП цилиндрической формы, с просветом в центре. Все процессы, начиная со связывании МП с ПЛ и кончая образованием П, являются Са2+-зависимыми.
3. Индукция апоптоза клеток-мишеней, опосредованная поверхностными рецепторами на их плазмолемме. Происходит в результате Са2+-независимого взаимодействия антигена Fas (АРО-1, CD95) на плазмолемме клеток-мишеней с Fas-L (Fcis-лигандом - литическим эффектором из семейства ФИО) на поверхности Тк. Механизмы передачи сигнала с поверхности клетки на систему протеолитических ферментов семейства ICE, реализующих программу апоптоза, остаются неясными.
Взаимодействие Fas и Fas-L способствует также регуляции численности популяции самих Т-лимфоцитов. Активированные Т-лимфоциты резко усиливают выработку и экспрессию белка Fas (сначала неактивного) и Fas-L. По прошествии нескольких дней Fas активируется, связывается с Fas-L на этих же или других активировагнных лимфоцитах, вызывая запуск программы их апоптоза. Тем самым ограничивается срок существования активированных Тк, что предотвращает возможность избыточных повреждений тканей и развития хроническог о воспаления.
Гуморальное токсическое воздействие Тк может, наряду с описанными выше контактными взаимодействиями, вызывать гибель клетокмишеней. Оно осуществляется путем секреции Тк токсических меди-
- 234 -
аторов - лимфотоксина (ФНОР), ФНОα, ИЛ-1, ИФНγ, повышения концентрации активных радикалов кислорода, простагландина Е2, подавления выработки рецепторов.
Т-клетки ГЗТ (Тгзт) являются эффекторными клетками иммунных реакций гиперчувствительности замедленного типа. Они обладают поверхностным антигеном CD4 и экспрессируют ТКР совместно с молекулами МНС II класса. Тгзт распознают чужеродные антигены, преимущественно продуцируемые внутриклеточными микроорганизмами. В последние годы накоплены данные, свидетельствующие о том, что роль Тгзт фактически исполняют Тх1, которые и обеспечивают течение указанных реакций.
Активация Тгзт сходна с таковой Тх (происходит в результате взаимодействия с АПК) и сопровождается секрецией ИФНγ, (активирующею макрофаги), МИФ (фактора, ингибирующего миграцию), МХФ (хемотаксического фактора макрофагов), и других лимфокинов, которые воздействуют на макрофаги, клетки Лангерганса (а также другие АПК), Тк и нейтрофилы. Таким образом, Тгзт, в отличие от другого класса эффекторов клеточного иммунитета (Тк) сами не обладают непосредственным цитотоксическнм действием, однако обеспечивают защитные реакции, активно вовлекая в них клетки других типов
.
Проявлением реакций ГЗТ служат очаги хронического воспаления, состоящие преимущественно из макрофагов и лимфоцитов, часто формирующих характерные компактные структуры - гранулемы. Их основу образуют скопления макрофагов (возникающие под действием МХФ и МИФ), которые находятся в активироваггном состоянии (эффект ИФНγ).
В-ЛИМФОЦИТЫ И ПЛАЗМАТИЧЕСКИЕ КЛЕТКИ
Функция В-лимфоцитов:
1)распознавание антигенов иммуноглобулиновыми рецепторами,
2)обеспечение реакций гуморального иммунитета,
3)дифференцировка в плазмоциты - клетки, вырабатывающие иммуноглобулины.
Развитие В-лимфоцитов в красном костном мозге
В-лимфоциты у птиц (у которых они впервые были выявлены) развиваются из клеток-предшественнпков в особом лимфоидном органе –
- 235 -
фабрициевой сумке (Buisa Fabricii), с чем и связано название этих клеток. У млекопитающих, в том числе у человека, В-лимфоциты развиваются из клетокпредшественников в красном костном мозге (у плода - первоначально в желточном мешке, печени и селезенке). В ходе пролиферации и дифференцировки в костном мозге у этих клеток в отсутствие антигенной стимуляции происходит реаранжировка части генома, ответственной за антигенную специфичность В-лимфоцитов. В р-зультате образуются клетки с огромным (порядка 109 вариантов) репертуаром антиген-распознающих рецепторов.
Взаимодействие В-лимфоцитов с антигенами и их участие в иммунных реакциях:
1.Из костного мозга наивные В-лимфоциты попадают в кровь, неся на
своей поверхности молекулы иммуноглобулинов (IgM), которые представляют собой специфические антиген-распознающие рецепторы (до 104-105/клетку), а также характерные маркеры CD19, CD20, CD21, CD22 и CD23. В-лимфоциты экспрессируют на плазмолемме молекулы МНС I и II классов, рецепторы к СЗкомпоненту комплемента и Fcучасткам молекул иммуноглобулинов.
2.Из крови, в которой В-клетки составляют 10-2.0% циркулирующих лимфоцитов, они направляются в периферические иммунные органы и заселяют их В-зависимые зоны. В последних в результате взаимодействия с антигенами (и Тх) происходит их активация и пролиферация. Она завершается созреванием
идифференцировкой активированных В-лимфоцитов в плазматические клетки, продуцирующие антитела, и В-клетки памяти. Антитела вырабатываются и самими активированными В-лимфоцитами, однако их основным источником в организме служат плазматические клетки.
3.Часть активированных В-лимфоцитов с током лимфы возвращается в кровь, а из нее попадает в различные органы (в особенности, в очага воспаления), где они превращаются в плазматические клетки.
Роль В-лимфоцитов в местной иммунной системе слизистых оболочек. Значительная доля активированных В-лимфоцитов направляется в слизистые оболочки, в особенности, в те, которые послужили входными воротами для данного антигена. Слизистые оболочки всех систем организма содержат значительное количество лимфоидной ткани, которая обозначается термином MALT (аббревиатура англ. Mucosa-Associated Lymphoid Tissue -
ассоциированная со слизистыми обо-
- 236 -
лочками лимфоидная ткань) и образует основу местной иммунной системы слизистых оболочек. Здесь активированные В-лимфоциты дифференцируются в плазматические клетки, в том числе вырабатывающие секреторные IgA, которые обеспечивают местный гуморальный иммунитет слизистых оболочек, защищающий их от колонизации микроорганизмами и от возможного внедрения микробов в ткани.
Активация В-лимфоцитов происходит под влиянием двух сигналов:
1)антигена, связывающегося со специфическим иммуноглобулиновым рецептором на поверхности В-лимфоцита;
2)контактного взаимодействия В-лимфоцита с Тх, секретируюшим ряд интерлейкинов. Это взаимодействие Тх осуществляют лишь с собственными В- лимфоцитами, обладающими соответствующими детерминантами МНС, т.е. оно является МНС-рестриктированным. Необходимость во втором сигнале отсутствует при воздействии тимус-независимых антигенов (см. ниже).
Механизм активации В-лимфоцитов включает последовательность явлений, сходную с наблюдаемой при активации Т-лимфоцитов (см. выше) и завершается усилением экспрессии ряда генов, в частности, связанных с пролиферацией клеток и их дифференцировкой в плазматические клетки - антителопродуценты.
Взаимодействие В-лимфоцитов с антигенами протекает неодинаково и зависит от природы антигенов, что позволило разделить последние на тимусзависимые и тимус-независимые.
1.Тимус-зависимые антигены неспособны активировать В-лимфоциты в отсутствие второго сигнала, обусловленного Тх, что послужило основанием к их наименованию. К этой группе относится большинство существующих антигенов.
2.Тимус-независимые антигены могут эффективно стимулировать В- лимфоциты (вызывая их активацию с последующей пролиферацией и дифференцировкой) без участия второго сигнала. В эту группу входит лишь небольшое число антигенов с многократно повторяющимися эпитопами, которые перекрестно связывают мембранные иммуноглобулиновые рецепторы В-лимфоцитов. К таким антигенам относятся, например, высокомолекулярные полисахариды микроорганизмов.
Взаимодействие В-лимфоцитов с Тх
1. Антиген-распознающие рецепторы В-лимфоцита специфически связывают антиген, который далее поглощается механизмом рецептор-
- 237 -
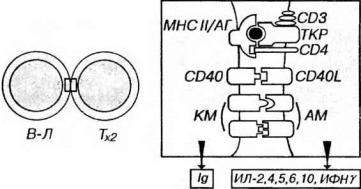
но-опосредованного эндоцитоза, подвергается процессингу и экспрессируется на поверхности В-лимфоцита в виде пептидов, связанных с молекулами МНС II класса (рис. 8-7).
2.Тх, ранее активированный данным антигеном в ходе взаимодействия с АПК, распознает комплекс молекулы МНС II класса/антиген на поверхности В- лимфонита. При этом в Тх происходит внутренняя перестройка, отчасти сходная с наблюдаемой в Тк при контакте с клеткой-мишеныо - ориентация его органелл в направлении В-клетки. Благодаря этому, очевидно, Тх способен осуществлять направленную секрецию интерлейкинов (главным образом, ИЛ-2, -4, -5, -6, -10) и ИФНγ на поверхность В-лимфоцита.
3.Указанные вещества активируют В-лимфоциты, стимулируют их пролиферацию и дифференцировку в плазматические клетки, способствуют переключению класса вырабатываемых антител. Активацию В-лимфоцита обеспечивают и ею непосредственные контактные взаимодействия с Тх, включающие связывание рецептора СD40 на плазмолемме В-лимфоцита с его лигандом (СD40L), экспрессируемым на поверхности активированною Тх. Взаимодействие СD40-СD40L необходимо для последующего переключения клетки с синтеза IgМ на продукцию иммуноглобулинов других изотопов IgG и IgA.
Рис. 8-7. Взаимодействие В-лимфоцита с Тх. Антиген (АГ) поглощается В- лимфоцитом, подвергается процессингу и экспрессируется на его поверхности в комплексе с молекулами МНС II класса (МНС II). Этот комплекс распознается Тх2 (посредством TKP и CD4), который секретирует ряд интерлейкинов на поверхность В- лимфоцита, активируя его, стимулируя пролиферацию и дифференцировку в плазматическую клетку, продуцирующую иммуноглобулины. Активации В-лимфоцита способствует контактное взаимодействие рецептора CD40 на его плазмолемме с лигандом CD40L на поверхности активированного Тх. Эффективность кооперации клеток повышается в результате взаимодействия костимулирующих молекул (КМ) на поверхности В-лимфоцита с адгезионными молекулами (AM) на плазмолемме Тх2.
- 238 -
Первичный гуморальный иммунный ответ развивается при первой встрече с антигеном и вызывает выработку небольшого количества антител. По прошествии определенного времени уровень антител обычно существенно падает.
Вторичный гуморальный иммунный ответ возникает при повторном попадании антигена и характеризуется быстрым развитием и продукцией больших количеств антител. Он обеспечивается благодаря активности В- и Т- лимфоцитов памяти (Вп и Тп). Эти клетки образуются при начальном воздействии антигена.
В-клетка памяти (Вп) - долгоживущая клетка, обеспечивающая быструю пролиферацию и дифференцировку В-лимфоцитов в плазмоциты при повторном контакте с антигенами. В отличие от виргильных лимфоцитов, которые в периферических иммунных органах живут всего несколько дней и погибают, если не встречаются со своим специфическим антигеном, Вп могут жить в течение нескольких месяцев и даже лет, не делясь и участвуя в рециркуляции.
ПЛАЗМАТИЧЕСКИЕ КЛЕТКИ
Плазматическая клетка (плазмоцит) - неподвижная или очень слабо подвижная, короткоживущая (2-3 сут, по другим сведениям - до 10-30 сут.) клетка - конечный этап развития В-лимфоцита. В ходе дифференцировки из В- лимфоцита она утрачивает рецепторы к СЗ-компоненту комплемента, мембранные иммуноглобулины и молекулы МНС, а также маркеры СD19 и
СD21.
Функциональные свойства плазматических клеток. Функция плазматических клеток заключается в обеспечении гуморального иммунитета путем выработки антител. За 1 секунду каждый плазмоцит синтезирует до нескольких тысяч молекул иммуноглобулинов (более 10 млн. молекул в час). Продуцируемые иммуноглобулины относятся к пяти классам (см. ниже), причем плазматические клетки способны переключаться с выработки иммуноглобулинов одного класса на другой.
Переключение классов продуцируемых иммуноглобулинов (переключение изотипов) происходит в развивающихся плазматических клетках примерно с 1- суточным интервалом - с IgM на IgG или IgА - без изменения их антигенсвязывающего участка. Процесс переключения связан с обратимыми изменениями процессинга транскриптов РНК. а также с необратимой рекомбинацией соответствующих участков ДНК. Одна клетка может синтезировать до трех классов иммуно-
- 239 -
глобулинов одновременно. Процесс переключения классов иммуноглобулинов контролируется цитокинами. Описаны иммунодефицитные состояния, связанные с нарушением переключения изотопа IgМ на IgG или (см. ниже).
Созревание аффинности продуцируемых иммуноглобулинов - выработка вновь образующимися плазмоцитами антител с возрастающей аффинностью (сродством) к данному антигену. Это явление служит отражением процессов соматических гипермутаций и селекции активированных В-лимфоцитов с высокоаффинными рецепторами, которые происходят в периферических органах кроветворения и иммуногенеза при продолжительном антигенном воздействии (см. главу 9).
Распределение плазматических клеток в организме. В норме плазматические клетки не циркулируют в крови. Они располагаются в красном костном мозге (составляя 1-2% его клеток), лимфатических узлах (преимущественно в мозговых тяжах и герминативных центрах), в белой пульпе селезенки. Значительное количество этих клеток характерно для рыхлой волокнистой соединительной ткани (см. главу 10), в особенности, образующей собственную пластинку слизистых оболочек (см. выше) и строму различных желез - например, слезных, слюнных, молочных (где преобладают клетки, продуцирующие IgA).
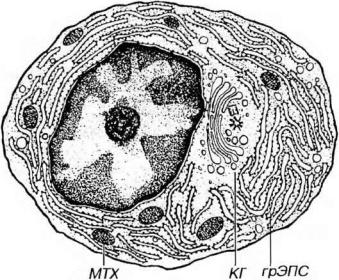
Рис. 8-8. Ультраструктурная организация плазматической клетки. КГ - комплекс Гольджи, МТХ - митохондрии.
- 240 -