

Bykov-_gistologia_obschaya
.pdfЭозинофилия (повышенное содержание эозинофилов в крови) наиболее выражена при аллергических состояниях (бронхиальной астме, аллергическом рините, аллергическом дерматите, пищевой аллергии), когда содержание эозинофилов увеличивается в несколько раз. Она характерна также для паразитарных заболеваний (при которых ее добавочному усилению способствует свойственный им аллегический компо-
нент и достигая у отдельных больных 90% общего числа лейкоцитов), физиологическая эозннофнлия свойственна первым трем месяцам жизни.
Эозинопения (сниженное содержание эозинофилов в крови) отмечается при острых инфекциях, введении глюкокортикоидов, АКТГ.
Размеры эозинофильных гранулоцитов на мазках больше, чем нейтрофильных и составляют 12-17 мкм.
Форма эозинофилов на мазках и в тканях - окрутлая, иногда с небольшими выпячиваниями (псевдоподиями). В мокроте и носовой слизи встречаются эозинофилы в виде отростчатых "клеток-медуз". Плазмолемма содержит низкоаффинные рецепторы к IgG, компонентам комплемента, высокоаффинные рецепторы к IgЕ (последние отсутствуют у нейтрофилов), цитокинам, гормонам, а также адгезивные молекулы.
Ядра эозинофильных гранулоцитов обычно сегментированные (состоят из двух, реже трех сегментов), светлее (содержат меньше гетерохроматина), чем ядра нейтрофилов. Изредка могут встречаться палочкоядерные и юные формы, отдельный подсчет которых обычно не производится.
Рис 7-11. Эозинофильный гранулоцит. 1 - ультраструктурная организация, 2 - эозинофильная гранула (ЭГ). АГ - азурофильная гранула, К - кристаллоид.
- 201 -
Цитоплазма эозинофильных гранулоцитов содержит умеренно развитые органеллы, многочисленные пузырьки, элементы цитоскелета, включения гликогена, липидные капли и гранулы двух основных шипов (рис. 7- 11). Предполагается также наличие особого третьего типа мелких гранул (микрогранул).
1. Специфические (эозинофильные) гранулы - наиболее характерный признак эозинофильных гранулоцитов; содержатся в количестве около 200 гранул на клетку (составляя более 95% всех гранул). Они окружены мембраной, имеют овальную или полигональную форму, крупные размеры (0.5-1.5х0.2-1.0 мкм), различную (чаше всего - среднюю) электронную плотность. Зрелые гранулы в большинстве содержат плотные кристаллоидные структуры, расположенные по их длине и погруженные в менее электронно-плотный мелкозернистый матрикс. Эти кристаллоиды имеют белковую природу и характеризуются кубической решеткой с периодом около 4 нм. Так как в эозинофильных гранулах находится ряд гидролитических ферментов, их рассматривают как видоизмененные лизосомы.
Содержимое специфических гранул:
(2) Главный основной белок (МBР, от англ. Major Basic Protein; название отражает высокое содержание этого белка, составляющего 50% общего белка специфических гранул, и его основную реакцию) образует их кристаллоид и обусловливает их эозинофилию. Содержит высокие концентрации аргинина, обладает мощным антигелъминтным, антипротозойным и антибактериальным эффектами. Токсичен для клеток других тканей (в частности, для эпителия слизистых оболочек воздухоносных путей и пищеварительного тракта). Вызывает гиперреактивность гладких мышц в бронхах. Индуцирует дегрануляцию базофилов, тучных клеток и тромбоцитов, активирует нейтрофилы. Инактивирует гепарин, гистамин, простагландины.
(2) Другие белки специфических гранул располагаются в их матриксе. К
ним относятся:
- эозинофильный катионный белок - токсичен для бактерий, гельминтов, простейших и клеток организма хозяина;
- 202 -
-эозинофильная пероксидаза (отличается от миелопероксидазы нейтрофильных гранулоцитов) - обладает широким спектром антимикробной и антипаразитарной активности в присутствии перикиси водорода;
-эозинофильный нейротоксин - обладает противопаразитарным действием, токсичен для клеток нервной системы;
-гистаминаза - разрушает гистамин; продукты расщепления гистамина оказывают на эозинофилы хемотаксическое действие.
2. Азурофильные (неспецифические, первичные) гранулы -
немногочисленные (менее 5% всех гранул), крупные и средних размеров (0.1- 0.5 мкм), округлой формы, с плотным содержимым. Представляют собой лизосомы и содержат кислую фосфатазу, арилсульфатазу (инактивирует лейкотриены и присутствует в очень большом количестве) и другие ферменты. Содержание этих гранул снижается по мере созревания клетки.
Цитофизиология эозинофильных гранулоцитов
Участие эозинофилов в защите от бактерий, грибов, простейших и гельминтов. Эозинофилы, как и нейтрофилы, способны поглощать и подвергать внутриклеточному уничтожению бактериальные клетки и споры патогенных грибов. Активность фагоцитарного уничтожения микробов у эозинофилов при этом обычно ниже, чем у нейтрофилов. Вместе с тем, эозинофильные гранулоциты являются главными клеточными элементами, обеспечивающими высокоэффективную защиту организма от простейших и гельминтов. Они способны уничтожать паразитов непосредственно в кровеносном русле. Выселяясь из кровеносных сосудов, они направляются в слизистые оболочки, где обеспечивают уничтожение паразитов - как внедрившихся в ткани, так и находящихся в просвете органа (обычно кишки). Они окружают паразиты, вступают с ними в контакт, и, активируясь, осуществляют дегрануляцию - выбрасывают токсическое содержимое своих гранул, обладающее высокой противопаразитарной активностью и одновременно вызывающее приток других эффекторных клеток. Цитотоксический эффект в противопаразитарном иммунитете является антителозависимым: активации и прикреплению эозинофила к поверхности паразита способствует наличие на его плазмолемме рецепторов к 1gЕ, и компонентам комплемента.
Иммунорегуляторная функция эозинофильных гранулоцитов
обеспечивается в результате их поступления в зону иммунных реакций и ограничения ее распространенности. Они привлекаются в эту зону продуктами,
- 203 -
выделяющимися в ходе иммунных реакций, которые они подвергают инактивации, одновременно угнетая деятельность продуцирующих их клеток. Эта функция осуществляется благодаря способности эозинофилов нейтрализовать лейкотриены, захватывать иммунные комплексы, связывать и разрушать гистамин и угнетать дегрануляцию тучных клеток и базофилов. Фосфолипаза эозинофилов расщепляет ФАТ.
Вместе с тем, активированные эозинофилы сами вырабатывают ФАТ (являясь его главным источником в организме) и лейкотриены, которые вызывают увеличение проницаемости сосудов и сокращение гладких мышц. При дегрануляции эозинофилов выделяются продукты, токсические для тканей человека. Поэтому, наряду с защитой тканей от действия продуктов иммунных реакций, эозинофилы способствуют и их повреждению. Так, установлено, что они являются важным звеном в патогенезе бронхиальной астмы, в частности, играют существенную роль в повреждении бронхиального дерева и респираторного отдела легких, а также в поддержании бронхоспастического синдрома. Помимо участия в регуляции реакций острого и хронического воспаления, эозинофилы, вырабатывая ряд цитокинов (ГМ-КСФ, ИП-1, ИЛ-5, ФНОа), могут также играть определенную роль в регуляции различных процессов, в частности, роста опухолей и заживления ран.
МОНОЦИТЫ
Моноциты - самые крупные из лейкоцитов; относятся к агранулоцитам. Они образуются в красном костном мозге, откуда попадают в кровь, в которой находятся от 8 ч до 3-4 сут. и, по-видимому, дозревают. Общее число моноцитов в крови у взрослого составляет 1.7-2.0 х 109 клеток, из которых 3/4 находятся в пристеночном пуле. Из кровеносного русла моноциты перемещаются в ткани со скоростью 4-10 х 108 клеток/сут. Внесосудистый пул моноцитов почти в 20 раз превышает их количество в циркуляции. В тканях под влиянием микроокружения и стимулирующих факторов они превращаются в различные виды макрофагов. Моноциты в совокупности с макрофагами образуют единую моноцитарно-макрофагальную систему или систему мононуклеарных фагоцитов (последнее название произошло от традиционного подразделения всех фагоцитов на полиморфноядерные (сегментоядерные), то есть нейтрофилы, и мононуклеарные (с несешентированным ядром), то есть моноциты.
Функции моноцитов в значительной мере связаны с их превращением в макрофаги после миграции из сосудов в ткани, хотя частично они могут реализовываться и самими моноцитами еще до этого превращения. К ним относятся:
- 204 -
1)Обеспечение реакций неспецифической защиты организма против микробов, опухолевых и зараженных вирусами клеток;
2)Участие в специфических (иммунных) защитных реакциях - в составе как их афферентного звена (в качестве антиген-представля-
юших клеток), так и эфферентного звена (в качестве эффекторных клеток);
3)Захват и внутриклеточное переваривание различных стареющих и погибших клеток и постклеточных структур (в том числе форменных элементов крови), а также их фрагментов; обеспечение метаболической переработки и реутилизации продуктов их распада (например, железа гемоглобина разрушенных эритроцитов).
4)Секреция различных веществ, которые регулируют: (а) состояние межклеточного вещества (лизосомальные протеазы, коллагеназы, эластазы, активатор плазминогена и др.); (б) функциональную активность и пролиферацию клеток других типов (монокины - разновидность цитокинов, выделяемых моноцитами/макрофагами).
Содержание моноцитов в крови взрослого в норме: абсолютное - 240-700
клеток/мкл, относительное - 6-8%; у детей в течение 1-й недели жизни - 10-20%.
Моноцитоз (повышенное содержание моноцитов в крови) наиболее часто служит проявлением воспалительных или опухолевых заболеваний, а также системных заболеваний крови.
Моноцитопения (сниженное содержание моноцитов в крови) в качестве изолированного состояния встречется редко. Содержание моноцитов снижено при ряде заболеваний системы крови - анапластических анемиях, некоторых лейкозах; оно надает после введения глюкокортикоидов.
Размеры моноцитов на мазках - 18-20 мкм. Они являются самыми крупными клетками среди лейкоцитов (см. рис. 7-1).
Форма моноцитов на мазках - округлая, под электронным микроскопом обнаруживаются различные цитоплазматические выпячивания.
Ядро моноцитов - крупное (занимает до половины площади клетей на мазке), эксцентрично расположенное, бобовидной или подковообразной формы (реже - дольчатое), светлое (хроматин рассеян в виде мелких гранул), с одним или несколькими мелкими ядрышками.
- 205 -
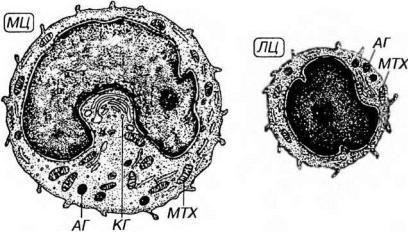
Цитоплазма моноцитов - слабобазофилъная, содержит многочисленные мелкие митохондрии, короткие цистерны грЭПС, вариабельное число свободных рибосом, полисом, сравнительно крупный комплекс Гольджи (рис. 7-12). Цитоскелет моноцитов хорошо развит; множественные микрофиламенты, концентрирующиеся в периферических участках его цитоплазмы под плазмолеммой в области формирующихся
псевдоподий обеспечивают его активные амебоидные движения. В цитоплазме присутствуют азурофильные гранулы (лизосомы), сходные с таковыми в нейтрофилах и богатые гидролитическими ферментами.
Рис. 7-12. Ультраструктурная организация моноцита (МЦ) и лимфоцита (ЛЦ). АГ - азурофильные гранулы, КГ - комплекс Гольджи, МТХ - митохондрии.
Антимикробные системы моноцита включают лизоцим, лактоферрин,
кислую фосфатазу, арилсульфатазу, катионные белки, миелопероксидазу, перекись водорода и другие биоокислители, а также токсический метаболит - окись азота (NO), которая синтезируется в цитоплазме при их активации.
Цитофизиология моноцитов и их роль в системе мононуклеарных фагоцитов
Моноциты активно выселяются в ткани из сосудистого русла, причем эта миграция усиливается под влиянием продуктов, выделяемых поврежденными тканями, микробами, а также под действием цитокинов. Моноциты обладают высокой активностью фагоцитоза и способны осуществлять иммунный фагоцитоз благодаря взаимодействию их плазмолеммы с опсонизированными микроорганизмами, которое опосредуется рецепторами к IgG и СЗ-компоненту комплемента. При фагоцитозе в моноцитах, как и в нейтрофилах, генерируются
- 206 -
токсические биоокислители (перекись водорода, супероксидный и гидроксильный радикалы, синглетный кислород), а также окись азота. Моноциты, как и образующиеся из них макрофага, способны также к нефагоцитарному уничтожению микрорганизмов, путем воздействия на них микробоцидных веществ, секретируемых в межклеточное пространство.
Превращение моноцитов в макрофаги происходит в тканях под влиянием местных факторов микроокружения. Некоторые исследователи полагают, что до этого превращения моноциты способны несколько раз делиться. Образующиеся макрофаги обладают, наряду с общими свойствами, некоторыми частными отличиями, обусловленными ткане- и органоспецифическими особенностями их существования и функционирования.
Моноциты, мигрирующие в ткани, дают начало макрофагам соединительной ткани (гистиоцитам), ряду органоспецифических макрофагов - клеткам Купфера печени, альвеолярным макрофагам легкого, макрофагам костного мозга, селезенки, тимуса, лимфатических узлов, полостей тела (перитонеальным, плевральным, перикардиальным), центральной нервной системы (микроглии), остеокластам (рис. 7-13). Предполагают, что и специализированные макрофаги в тканях способны к делению, однако оно недостаточно для поддержания их популяций, которое осуществляется путем постоянного притока моноцитов из крови и их преобразования в макрофаги.
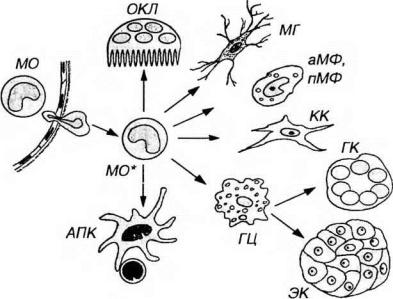
Рис. 7-13. Основные направления преобразования моноцитов в различные типы макрофагов (МФ) и антиген-представляющие клетки (АПК). МО - моноцит (в
- 207 -
просвете кровеносного сосуда и мигрирующий через его стенку); МО* - моноцит в тканях, дифференцирующийся в АПК или один из видов МФ: гистиоцит (ГЦ), клетку Купфера (КК) синусодов печени, альвеолярный МФ (аМФ), перитонеальный МФ (пМФ), клетку микроглии (МГ) и остеокласт (ОКП). В очаге воспаления ГЦ могут дать начало гигантской клетке (ГК) или эпителиоидным клеткам (ЭК).
Структурно-функциональные изменения моноцитов при их превращении в макрофаги включают:
1)существенное увеличение размеров клетки (до 25-50 мкм), а также содержания в ее цитоплазме митохондрий, пиноцитозных пузырьков и, в особенности, лизосом, размеров комплекса Голъджи;
2)преобразования плазмолеммы с формированием значительного числа складок, увеличением количества микроворсинок, нарастанием содержания рецепторов к IgG и СЗ-компоненту комплемента;
3)повышение активности дыхательных и лизосомальных ферментов,
одновременное снижение содержания пероксидазы;
4)усиление подвижности, общей метаболической активности, адгезивных свойств, способности к пиноцитозу и фагоцитозу, общее возрастание микробицидности;
5)изменения чувствительности к гормонам.
Фагоцитоз у макрофагов, как у моноцитов и нейтрофилов, сопровождается "респираторным взрывом". Он может осуществляться как неиммунный фагоцитоз (в отсутствие воздействия специфических факторов сыворотки) или как иммунный фагоцитоз (после опсонизации, благодаря наличию рецепторов к IgG и СЗ-компоненту комплемента на плазмолемме макрофага.
Макрофаги из различных органов и тканей обладают неодинаковыми свойствами, в частности, различиями в способности к уничтожению микробов; определенная специфика характерна и для отдельных клеток среди однотипных макрофагов.
Резистентность микроорганизмов к действию микробицидных механизмов макрофагов обеспечивается несколькими путями. Так, некоторые микробы, например, возбудители туберкулеза (Mycobacterium tuberculosi) и токсоплазмоза (Toxoplasma gondii), избегают действия микробицидных механизмов и выживают в фагосомах макрофагов благодаря тому, что выделяют вещества, которые препятствуют слиянию лизосом с фагосомами. Другие (например, лейшмании) сохраняют жизнеспособность в фаголизосомах,
- 208 -
так как обладают стенкой, резистентной к действию лизосомальных ферментов и низких значений pH, третьи (например, Trypanosoma cruzei) могут проникать из фагосом в гиалоплазму.
Активация макрофагов - процесс, обеспечивающий дальнейшее усиление их метаболической, локомоторной, фагоцитарной, антимикробной, противоопухолевой и секреторной способности - происходит при непосредственном контакте с микроорганизмами, а также под вли-
янием их продуктов или цитокинов - ИЛ-2, ИЛ-4, ФНО, ИФНγ, ФАТ, колониестимулирующих факторов (КСФ). Активированные макрофаги приобретают способность к уничтоженшо ряда микроорганизмов, которые могли выживать в фаголизосомах неактивированных клеток и даже разрушать их. Индукция выработки значительных количеств окиси азота усиливает цитотоксичность макрофагов по отношению к опухолевым клеткам и микроорганизмам. Макрофаги, неспособные уничтожить фагоцитированные ими микроорганизмы, благодаря своей подвижности превращаются в их разносчиков и тем самым могут содействовать распространению инфекции по организму.
Секреция активированных макрофагов. При активации усиливается продукция макрофагами и секреция ими во внеклеточное пространство различных веществ - ИЛ-1, ФНОα, ТРФР, простагландинов, лейкотриенов, ФАТ, ТФРα, ТФРβ, компонентов комплемента, ИФН, катионных белков, свободных радикалов кислорода, перекиси водорода, окиси азота, хемотаксических факторов для нейтрофилов, М-КСФ, ГМ-КСФ. Часть этих веществ важна для непосредственной защиты от микроорганизмов и опухолевых клеток, другая - оказывает влияние на сами макрофаги и другие клетки, регулируя активность воспалительных реакций. Так, ИЛ-1, ФНОα и ряд других пептидов, воздействуя на гипоталамический терморегуляторный центр, играют роль эндогенных пирогенов (от греч. pyros - огонь и genes - происшедший), т.е. веществ, вызывающих повышение температуры тела. Микробные продукты, обусловливающие выделение эндогенных пирогенов, носят название экзогенных пирогенов. Секретируемьге активированными макрофагами лизосомальиьге фермента в сочетании с микробицидными веществами обеспечивают уничтожение микроорганизмов нефагоцитарньгм путем, однако они способны вызвать разрушение и окружающих их тканей. Секреция медиаторов воспаления макрофагами угнетается кортикостероидными препаратами.
Макрофаги обладают и другими механизмами влияния на воспалительные реакции, так как они способны разрушать компонента комплемента, иммуноглобулины, кинины. В очагах воспаления макрофаги могут стимулировать процессы регенерации ткани путем удаления погибших клеток и
- 209 -
секреции факторов, вызывающих пролиферацию и функциональную активацию фибробластов - клеток, обеспечивающих выработку компонентов межклеточного вещества.
Видоизменения макрофагов в тканях и особые виды макрофагов
В тканях могут встречаться макрофаги, перегруженные продуктами неполного переваривания фагоцитированных ими субстратов,
а также макрофаги, изменившиеся в результате взаимодействия между собой и с другими клетками, в первую очередь, лимфоцитами (в очагах хронического воспаления). Такие макрофаги приобретают ряд морфологических особенностей, столь характерных, что они служат их диагностическими признаками и обусловливают особые названия этих клеток:
1)"Пылевые" клетки - альвеолярные макрофаги легкого, перегруженные частицами пыли из вдыхаемого воздуха и выявляемые в мокроте;
2)Клетки ”сердечных пороков" - альвеолярные макрофаги, содержащие в цитоплазме большое количество железа в результате переваривания эритроцитов, попадающих в просвет альвеол при некоторых пороках сердца вследствие повышения давления в легочных сосудах и увеличения проницаемости их стенки.
3)”Пенистые”, или "ксантомные" (от греч. xantos - желтый - по цвету включений) клетки - макрофаги, с резко вакуолизированной цитоплазмой, перегруженные различными по химическому составу липидами (например, в очагах атеросклеротических поражений артерий, при повышенных уровнях липидов в крови, при наследственных заболеваниях, связанных с накоплением липидов - болезни Нимана-Пика, Гоше, Фабри и др.).
4)Гигантские многоядерные клетки - образуются в очагах хронического воспаления в результате слияния нескольких макрофагов друг с другом (см. рис. 7-13), поэтому их точнее следовало бы отнести к симпластам. Имеют разнообразную форму и нередко достигают очень крупных размеров, оправдывая свое название. В их цитоплазме могут находиться фагоцитированные микроорганизмы, различные клетки и их фрагменты.
5)Эпителиоидные клетки - располагаются в очагах хронического воспаления в виде рядов и скоплений, внешне напоминая клетки эпителия (что обусловило их наименование) - см. рис. 7-13. Характеризуются редукцией лизосомального аппарата, падением фагоцитарной активности при одновременном развитии синтетического аппарата. Специализируются на секреции различных регуляторных веществ (цитокинов, хемотаксических веществ, факторов роста) и ферментов в межклеточное пространство. Характер секретируемых веществ зависит от особенносей микроокружения этих клеток. Тем самым они оказывают сложное регулирующее действие на течение хронического воспаления.
-210 -