

Bykov-_gistologia_obschaya
.pdfновлению кроветворения. Общая масса миелоидной ткани в организме взрослого человека составляет 1.5-2 кг, снижаясь с возрастом. У пожилых людей эта ткань частично замещается жировой и в плоских костях.
Миелоидная ткань содержит СКК и является местом образования эритроцитов, гранулоцитов, моноцитов, тромбоцитов, В-лимфоцитов, предшественников Т-лимфоцитов и NK-клеток, в ней образуются также предшественники некоторых клеток соединительной ткани. В организме взрослого человека миелоидная ткань за сутки продуцирует и выделяет в кровь 2x1011 эритроцитов, 1010- 1011 гранулоцитов и 4x1011 тромбоцитов.
Лимфоидная ткань располагается в лимфоидных органах (органах иммунной системы) - тимусе, селезенке, лимфатических узлах, миндалинах, пейеровых бляшках, червеобразном отростке и многочисленных лимфоидных образованиях, имеющихся в стенке органов различных систем. Ее общая масса в организме взрослого человека достигает 1.5-2 кг, а количество входящих в ее состав лимфоцитов составляет 5-10х1011. В ней происходит образование Т- и В- лимфоцитов, а также плазматических клеток (конечной стадии дифференцировки В-лимфоцитов), которые взаимодействуя между собой, а также с макрофагами, дендритными антиген-представляющими и другими клетками, обеспечивают развитие и течение иммунных реакций. Характерной особенностью лимфоидной ткани служит то, что значительная часть образовавшихся в ней лимфоцитов погибает механизмом апоптоза в результате процессов селекции, связанных с отбором клеток, несущих необходимые рецепторы.
ОБЩИЕ ЗАКОНОМЕРНОСТИ РАЗВИТИЯ ФОРМЕННЫХ ЭЛЕМЕНТОВ КРОВИ
Источники развития форменных элементов крови и теории кроветворения. В настоящее время доказано, что общим источником развития всех форменных элементов крови служит плюрипотентная стволовая клетка крови (СКК). Это положение впервые сформулировано профессором А.А.Максимовым в начале нашего века в разработанной им унитарной теории кроветворения, которой противопоставляли ряд других теорий, допускавших развитие различных форменных элементов из двух, трех или большего числа отдельных стволовых клеток (дуалистическая, триалистическая и полифилетическая теории).
- 251 -
СТВОЛОВЫЕ КЛЕТКИ КРОВИ
Плюрипотентные стволовые клетки крови по строению напоминают малые лимфоциты, но могут быть идентифицированы иммуноцитохимически по набору антигенов на клеточной поверхности. Наиболее типичным маркерным признаком служит сочетание CD34+CD38-.
Локализация СКК. СКК сосредоточены у взрослого человека преимущественно в красном костном мозге, однако обнаруживаются в крови, циркулируя в которой они попадают в другие органы кроветворения. Очевидно, что СКК обладают уникальной для малодифференцированных клеток способностью выселяться из костного мола и, подобно зрелым форменным элементам, мигрировать через эндотелий венозных синусов в кровь. В красном костном мозге их содержание невелико (одна СКК приходится примерно на 2000 клеток), в циркулирующей крови СКК составляют 0.0001% от общего числа лейкоцитов (что соответствует соотношению 1 СКК : 1 млн. лейкоцитов). Важным источником получения СКК является плацентарная (пуповинная) кровь - концентрация СКК в ней достигает 0.16% от числа лейкоцитов, т.е. в 2-3 раза превосходит таковую в красном костном мозге взрослого. По способности к пролиферации СКК пуновинной крови в 10 раз превышают СКК костного мозга.
Основные свойства плюрипотентных стволовых клеток крови:
1.Обладают способностью к самоподдержанию без притока клеток извне
(т.е. к образованию в результате деления дочерних клеток, не отличающихся своим практически неограниченным пролиферативным потенциалом от родительской). Согласно иным взглядам, они обладают высоким, но все же ограниченным пролиферативным потенциалом. В соответствии с такими представлениями, гемипоэз обеспечивается благодаря тому, что в течение жизни индивидуума происходит последовательная смена одних клонов стволовых клеток другими (теория клональной последовательности, или каскадная теория).
2.Редко делятся (основное состояние - покоя); однако могут быть вовлечены в пролиферацию при значительных кровопотерях и при воздействии факторов роста. Деление СКК стимулируется фактором стволовых клеток (ФСК), который вырабатывается стромальными клетками костного мозга и фиксируется на поверхности стволовых клеток протоонкогенным белком с-кit.
Деление СКК может осуществляться в трех вариантах: (1) симметрично с образованием двух дочерних клеток, идентичных родительской (см. выше), (2) симметрично с появлением двух сходным образом ком-
- 252 -
митированных полустволовых клеток, (3) асимметрично путем так называемого квантального митоза с образованием одной стволовой и одной коммитированной полустволовой клетки.
3.Способны образовывать все виды форменных элементов крови (т.е. обладают истинной плюрипотентностью).
4.Устойчивы к действию повреждающих факторов (но сравнению с более дифференцированными клетками).
5.Располагаются в местах, хорошо защищенных от внешних воздействий (ячейки в костной ткани) и обладающих обильным кровоснабжением.
6.Циркулируют в крови, мигрируя в другие органы кроветворения.
КОММИТИРОВАНИЕ, ДЕТЕРМИНАЦИЯ И ДИФФЕРЕНЦИРОВКА КРОВЕТВОРНЫХ КЛЕТОК
В ходе гемопоэза плюрипотентные СКК делятся и дают начало клеткам, пролиферация которых приводит ко все большему ограничению направлений их развития. Этот процесс обусловлен последовательным коммитированием (ограничением потенций развития) и детерминацией (выбором направления развития) кроветворных клеток, которые на определенных этапах сопровождаются приобретением ими специфических структурных и функциональных признаков (дифференцировкой). Указанные процессы обусловлены внутренней программой развития гемопоэптческих клеток, которая реализуется лишь в условиях строго определенного для каждого типа клеток микроокружения - совокупности разнообразных физико-химических и трофических факторов, влияния гемопоэтинов (цитокинов, колониестимулирующих факторов - КСФ), контактных взаимодействий с другими гемопоэтическими и стромальными клетками, а также компонентами межклеточного вещества и др.
Гемопоэтические факторы роста (гемопоэтчны) вырабатываются стромальными компонентами кроветворных тканей и органов, в первую очередь, ретикулярными клетками. Они продуцируются также эпителиальными клетками тимуса, макрофагами, Т-лимфоцитами, жировыми клетками, клетками эндотелия, а также клетками, расположенными вне кроветворных тканей (например, эритропоэтин вырабатывается
- 253 -
клетками почек и печени). Они выделяются в кровь и могут действовать (а) дистантно (подобно гормонам) или (б) локально. Эффект гемопоэтинов проявляется в низких концентрациях, опосредован их связыванием со специфическими рецепторами на плазмолемме развивающихся клеток крови и заключается во влиянии на выживание, пролиферацию и дифференцировку этих
клеток.
Каждый этап развития конкретной линии клеток требует присутствия определенной комбинации гемопоэтинов. Отдельный гемопоэтический фактор может оказывать влияние па один или несколько типов развиваюишхся клеток.
Универсальным гемопоэтином служит ИЛ-3 (мульти-КСФ), который оказывает влияние как на самые ранние, так и на сравнительно поздние стадии гемопоэза. Основные эффекты ГМ-КСФ, Г-КСФ и М-КСФ связаны с их действием на родоначальные клетки, проявляющемся общей или раздельной стимуляций развили клеток линии гранулоцитов (нейтрофильных) и макрофагов. ИЛ-7 влияет на родоначальные клетки лимфоцитопоэза, стимулируя образование Т- и В-лимфоцитов.
КЛАССИФИКАЦИЯ КРОВЕТВОРНЫХ КЛЕТОК
На основании способности с самообновлению, клеточному делению и образованию форменных элементов различных типов кроветворные клетки можно разделить на шесть классов (рис. 9-2):
I класс - плюрипотентные стволовые клетки, которые могут образовывать любые форменные элементы и обладают способностью к самообновлению (см. выше).
II класс - частично детерминированные поли-, или мультипотентные родоначальные клетки (progenitor cells в англоязычной литературе), именуемые также полустволовыми клетками. Развиваются из СКК, способны к ограниченному самоподдержанию, являются полипотентными, однако прошедшими первый этап коммитирования клетками, т.е. дающими начало форменным элементам нескольких (но не всех) видов. Родоначальные клетки, относящиеся к данному и следующему (III) классам, называют также колониеобразующими единицами (КОЕ), поскольку в экспериментах на летально облученных мышах они способны давать колонии кроветворных клеток в их органах. Используемый иногда термин КОЕ-С (селезенки), применяется неоднозначно - одними авторами для обозначения клеток только I класса (СКК), а другими - также и полустволовых клеток (клеток II класса).
- 254 -
Частично детерминированные полипотентные родоначальные клетки включают родоначальную клетку лимфоцитоноэза (KOE-JI) и родоначальную клетку миелопоэза (КОЕ-ГЭММ), дающую начало гpaнулоцитам, эритроцитам, моноцитам и мегакариоцитам.
III класс - унипотентные (коммитированные) родоначальные клетки,
прошедшие новый этап коммитирования и детерминированные в направлении развития только одного вида форменных элементов (за исключением КОЕ-ГМ, дающей два вида - см. ниже). Они обладают низким потенциалом самоподдержания. Эти клетки не идентифицируются морфологически и внешне сходны с малыми лимфоцитами.
Унипотентные (коммитированные) родоначальные клетки включают: (1)
родоначальные клетки эритроцитов - БОЕ-Э - бурстобразуюшую единицу (происхождение термина см. ниже) и развивающуюся из нее КОЕ-Э, (2) КОЕМег - родоначальную клетку мегакариоцитов, (1) КОЕ-ГМ - родоначальную клетку гранулоцитов (нейтрофилышх) и моноцитов, дающую КОЕ-Г(Н) - родоначальную клетку гранулоцитов (нейтрофильных) и КОЕ-Мо - родоначальную клетку моноцитов, (2) КОЕ-Баз - родоначальную клетку базофилов, (5) КОЕ-Эо - родоначальную клетку эозинофилов, (6) коммитированные клетки лимфоцитопоэза - про-В-лимфоциты и протимоциты.
IV класс - морфологически распознаваемые предшественники - (precursors в англоязычной литературе) - бластные формы. Представляют отдельные линии развития форменных элементов. Пролиферативная активность этих клеток ограничена; способностью к самоподдержанию они не обладают. Название класса отражает тот факт, что, хотя морфологически все клетей этого класса сходны друг с другом, их можно идентифицировать при использовании стандартных гематологических методов окраски, не прибегая к выявлению иммуноцитохимических маркеров. Бластные формы имеют вид крупных клеток с базофильной цитоплазмой и светлым ядром, в котором хорошо определяются
ядрышки.
V класс - созревающие (дифференцирующиеся) клетки. Подвергаются структурной и функциональной дифференцировке, образуя соответствующий вид форменных элементов, в ходе которой они (за исключением лимфоцитов и моноцитов) утрачивают способность к делению.
VI класс - зрелые (дифференцированные) форменные элементы,
циркулирующие в крови. Неспособны к делению (за исключением лимфоцитов и моноцитов).
- 255 -
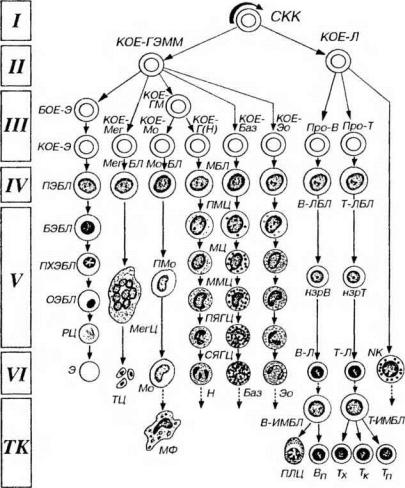
Рис 9-2. Схема кроветворения. I-VI - классы кроветворных клеток, СКК - стволовая клетка крови, КОЕ - колониеобразующая единица (родоначальная клетка): КОЕ-ГЭММ - КОЕ гранулоцитов, эритроцитов, моноцитов и мегакариоцитов, КОЕ-Э - КОЕ эритроцитов, КОЕ-Мег - КОЕ мегакариоцитов, КОЕ-ГМ - КОЕ гранулоцитов (нейтрофильных) и моноцитов, КОЕ-Г(Н) - КОЕ гранулоцитов (нейтрофильных), КОЕ-Мо - КОЕ моноцитов, КОЕ-Баз - КОЕ базофилов, КОЕ-Эо - КОЕ эозинофилов, КОЕ-Л - КОЕ лимфоцитопоэза, БОЕ-Э - бурст-образующая единица, про-В - про-В- лимфоцит, про-Т - про-Т- лимфоцит (протимоцит), ПЭБЛ - проэритробласт, БЭБЛ - базофильный эритробласт, ПХЭБЛ - полихроматофильный эритробласт, ОЭБЛ - оксифильный (ортохроматофильный) эритробласт, РЦ - ретикулоцит, Э - эритроцит. МегБЛ - мегакариобласт, МегЦ - мегакариоцит, ТЦ - тромбоциты, МоБЛ - моноцитобласт, ПМо - промоноцит, Мо - моноцит, МФ - макрофаг, МБЛ - миелобласты, ГШЦ - промиелоциты, МЦ - миелоциты, ММЦ - метамиелоциты, ПЯГЦ – палочкоядерные
- 256 -
гранулоциты, СЯГЦ - сегментоядерные гранулоциты (нейтрофильный - Н, базофильный - Баз, эозинофильный - Эо). В-ЛБЛ - В-лимфобласт, нзрВ - незрелый В-лимфоцит, В-Л - В-лимфоцит (зрелый), В-ИМБЛ - В-иммунобласт, ПЛЦ - плазмоцит, Вп - В-клетка памяти, Т-ЛБЛ - Т-лимфобласт , нзрТ - незрелый Т-лимфоцит, Т-Л - Т-лимфоцит (зрелый), Т-ИМБЛ - Т-иммунобласт, Тх - Т-хелпер, Тк - Т-киллер, Тп - Т-клетка памяти, ГТК - ЫК-клетка. Миграция зрелых клеток из крови в периферические ткани (ТК) обозначена пунктирными стрелками; пути рециркуляции лимфоцитов не отмечены.
Форменные элементы крови в тканях. Из зрелых форменных элементов лишь эритроциты и (частично) тромбоцита выполняют свои функции исключительно в кровотоке, лейкоциты же реализуют их после миграции в ткани. Часть клеток при этом подвергается дальнейшим преобразованиям (например, моноциты превращаются в макрофаги и дендритные АПК, лимфоциты под действием антигенной стимуляции подвергаются бласттрансформации и дальнейшей дифференцировке, основная часть В-лимфоцитов дифференцируется в плазматические клетки).
ЭРИТРОПОЭЗ
Эритропоэз (эритроцитопоэз) - процесс образования и созревания эритроцитов, происходящий в миелоидной ткани. Ход развития эритроцитов из стволовой клетки крови описывается последовательностью:
СКК -> КОЕ-ГЭММ -> БОЕ-Э -> КОЕ-Э -> проэритробласт > базофильный эритробласт -> полихроматофнльный эритробласт -> оксифильный (ортохроматофильный) эритробласт -> ретикулоцит -> эритроцит.
Эритрон - эритроидный дифферон, представляющий собой совокупность указанных форм - от эритроидных родоначальных клеток до зрелых эритроцитов (включая циркулирующие в крови).
БОЕ-Э и КОЕ-Э. БОЕ-Э - бурст-образующая единица (от англ. burst - взрыв) - названа так по своей способности быстро (взрывоподобно) образовывать на полутвердой среде колонию эритроидных клеток численностью в несколько сотен элементов. Она отличается от развивающейся из нее КОЕ-Э более высокой пролиферативной активностью, высокой чувствительностью к ИЛ-3 и низкой - к эритропоэтину.
Терминология, используемая для наименования дальнейших стадий развития клеток эритроидного ряда (следующих за КОЕ-Э) неоднозначна. Термины эритробласт, нормобласт и нормоцит применяются
- 257 -
разными авторами для обозначения одних и тех же клеточных форм. В настоящей книге использованы наименования указанных клеток, получившие наиболее широкое распространение в гематологической и гистологической литературе.
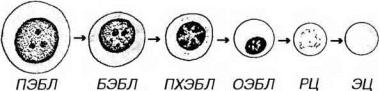
Процесс дифференцировки предшественников эритроцитов в зрелые форменные элементы включает (рис. 9-3):
(1)уменьшение размеров клетки;
(2)выработку и накопление гемоглобина в цитоплазме;
(3)постепенное снижение содержания и в конечном итоге утрату всех органелл;
(4)изменение окраски цитоплазмы от интенсивно базофильной (в связи с большим числом полирибосом) до оксифильной (обусловленной присутствием гемоглобина);
(5)снижение, а в дальнейшем (в конце стадии оксифильного эритробласта) - утрату способности к делению;
(6)конденсацию ядра и его последующее удаление из клетки.
Рис. 9-3. Эритропоэз. ПЭБЛ - проэритробласт, БЭБЛ - базофильный эритробласт, ПХЭБЛ - полихроматофильный эритробласт, ОЭБЛ - оксифильный (ортохроматофильный) эритробласт, РЦ - ретикулоцит, ЭЦ - эритроцит.
Проэритробласт - крупная клетка (диаметром около 18-22 мкм) с большим сферическим ядром, содержащим мелкодисперсный хроматин и дватри бледных ядрышка. Цитоплазма умеренно базофильна вследствие присутствия свободных рибосом. Клетка интенсивно пролиферирует, давая начало эритробластам. Последние развиваются в составе так называемых эритробластических островков.
Эритробластические островки - особые структурные комплексы в миелоидной ткани, обеспечивающие развитие эритробластов. Их центр образован телом макрофага, который своими многочисленными отростками охватывает окружающие его в один-два слоя эритробласты. По мере созревания эритробласты центробежно смещаются по длине отростков макрофага, удаляясь от его тела и отодвигаясь на периферию эритробластического островка, одновременно приближаясь к поверхности венозного синуса.
- 258 -
Базофилъный эритробласт - меньших размеров, чем проэритробласт (1216 мкм), с более мелким ядром, содержащим умеренно конденсированный хроматин и ядрышки. Цитоплазма резко базофильна благодаря высокому содержанию полисом, активно синтезирующих гемоглобин. Базофильные эритробласты активно делятся.
Полихроматофильный эритробласт характеризуется более мелкими размерами, чем батофильный эритробласт (10-12 мкм); его ядро более компактно, чем в базофильном эритробласте, глыбки хроматина в нем распределены в виде спиц колеса, ядрышко не выявляется. Цитоплазма окрашивается полихромно: она воспринимает как основные красители (вследствие наличия в ней многочисленных полисом), так и кислые (из-за накопления в ней оксифилыю окрашивающегося гемоглобина). Окраска цитоплазмы может быть либо диффузной и однородной, либо сочетать оксифилыше и базофильные участки. Скопления гемоглобина вокруг ядра часто имеют вид оксифильного перинуклеарного ободка. По мере накопления гемоглобина полисомы и другие органеллы редуцируются. Способность клетки к делению сохраняется.
Железо, необходимое для синтеза гема, поступает в цитоплазму эригробластов из двух источников: (1) непосредственно из крови (где оно связано с белком трансферрином) - путем транспорта, опосредованного рецепторами трансферрина на поверхности эригробластов; (2) из цитоплазмы макрофагов, контактирующих с эритробластами в эритробластических островках. Железо в составе ферритина (в комплексе с белком) выделяется на поверхность эригробластов в виде частиц диаметром 6 нм, которые связываются с их гликокаликсом и далее переносятся в их цитоплазму механизмом микропиноцитоза. Небольише скопления ферритина диаметром 0.1-0.3 мкм (сидеросомы) можно выявить в цитоплазме. Согласно расчетам, второй механизм переносит в 1000 раз больше железа, чем первый.
Оксифилъный (ортохроматофильный) эритробласт (нормобласт)
образуется путем дифференцировки из полихроматофильного эритробласта. По размерам он чуть крупнее эритроцита. Обладает оксифильно окрашенной цитоплазмой, богатой гемоглобином, в которой органеллы почти полностью отсутствуют. Ядро мелкое, компактное, пикнотическое, расположено эксцентрически. Способность к делению теряется.
Выталкивание ядра - наиболее важный этап в процессе превращения оксифильного эритробласта в ретикулоцит. Оно длится несколько минут и может происходить, когда клетка находится в составе
- 259 -
эритробластического островка или мигрирует через стенку кровеносных сосудов (синусов) костного мозга в кровоток. Этому процессу предшествует перестройка цитоскелета клетки, образующего структуру в виде манжетки, которая способствует активному выталкиванию ядра с тонким ободком окружающей его цитоплазмы за пределы клетки, где оно быстро фагоцитируется макрофагами.
Ретикулоцит представляет собой безъядерную (постклеточную) структуру. Его оксифильная цитоплазма, почти целиком заполненная гемоглобином, содержит остатки полирибосом и других органелл, которые выявляются при суправитальной окраске в виде базофильной сеточки. В кровотоке ретикулоцит в течение 24-48 ч превращается в зрелый эритроцит (см. главу 7).
Длительность всех этапов эритропоэза - от КОЕ-Э до образования зрелого эритроцита равна около 3-7 сут.
Регуляция процесса эритропоэза осуществляется рядом гуморальных факторов, из которых наибольшее значение имеют ИЛ-3 (стимулирует пролиферативную активность БОЕ-Э) и эритропоэтин (усиливает пролиферацию КОЕ-Э). Для нормального эритропоэза необходимы также железо, фолиевая кислота и витамин В12.
Эритропоэтин продуцируется у взрослого на 90% почкой, на 10% печенью (последняя, однако, служит главным его источником у плода) и вырабатывается в ответ на гипоксию. Его действие усиливается андрогенами, гормоном роста, тироксином и ослабляется эстрогенами (поэтому у женщин содержание эритроцитов и гемоглобина в крови ниже, чем у мужчин).
Недостаточная выработка эритропоэтина (например, при заболеваниях почек, некоторых эндокринных расстройствах - недостаточности гипофиза, щитовидной железы, коркового вещества надпочечника, мужском гипогонадизме) может вызывать развитие анемии, которая излечивается введением рекомбинантного эритропоэтина.
Применение эритропоэтина в качестве допинга у спортсменов для повышения физической работоспособности основано на увеличении переноса кислорода возросшим числом эритроцитов в крови. Последнее, однако, чревато риском развития тромботических осложнений из-за повышенной вязкости крови.
- 260 -