

Bykov-_gistologia_obschaya
.pdf(б) транс- (от лат. trans - по ту сторону), зрелую - вогнутой формы, обращенную к плазмолемме и связанную с отделяющимися от цистерн вакуолями. Между цистернами цис- и транс-поверхностей располагаются цистерны медиальной части комплекса Гольджи.
Транспорт веществ в комплексе Гольджи. Белки проникают в стопку цистерн комплекса Гольджи из транспортных пузырьков с цисповерхности, а выходят в вакуолях с транс-поверхности; каким образом осуществляется их перенос внутри комплекса, в ходе которого происходит их процессинг, остается неизвестным. Возможные пути этого транспорта описываются двумя моделями:
1)модель перемещения цистерн постулирует, что за счет слияния транспортных пузырьков на цис-поверхности непрерывно происходит новообразование цистерн (что легло в основу термина "формирующаяся поверхность"), в дальнейшем смещающихся к транс-поверхности, по достижении которой они распадаются на вакуоли ("зрелая поверхность"). Согласно этой модели, одни операции процессинга сменяются другими при перемещении самой цистерны по ходу изменений ее состава. Транспорт веществ из одной цистерны в другую, в соответствии с описанной моделью, отсутствует;
2)модель везикулярного транспорта предполагает, что цистерны не меняют своего расположения (остаются постоянно на своем месте), а продукты синтеза переносятся от цис- к транс-поверхности в пузырьках (везикулах), которые отпочковываются от предшествующей цистерны, сливаясь с последующей.
Функции комплекса Гольджи:
1)синтез полисахаридов и гликопротеинов (гликокаликса, слизи);
2)процессинг молекул: включение углеводных компонентов в гликопротеины, транспортируемые из грЭПС (терминальное гликозилирование), добавление фосфатных групп (фосфорилирование), жирных кислот (ацилирование), сульфатных остатков (сульфатирование), частичное расщепление белковых молекул (протеолитическая доработка). Каждый их указанных этапов процессинга веществ внутри комплекса Гольджи осуществляется в топографически определенном его компоненте (цис-, медиальных или транс-цистернах, а также сети транс-Голь- джи);
3)конденсация секреторного продукта (в конденсирующих вакуолях) и
образование секреторных гранул;
4)обеспечение новообразованных гранул мембраной (синтезированной в ЭПС) и упаковка в нее секреторных продуктов; в процессе секреции эта мембрана встраивается в плазмолемму, увеличивая площадь ее поверхности;
-51 -
5) сортировка белков на транс-поверхности (в сети транс-Голъджи) перед их окончательным транспортом. Направление последующего транспорта различных белков из комплекса Гольджи зависит от особенностей их гликозшшрования, фосфорилирования и сульфатирования. Сортировка производится посредством специфических мембранных рецепторных белков, которые распознают сигнальные участки на макромолекулах и направляют их в соответствующие пузырьки.
Транспорт белков из комплекса Гольджи осуществляется в составе трех важнейших потоков (рис. 3-10): (1) в гидролазные пузырьки (ранее называемые первичными лизосомами) - начально в виде окаймленных пузырьков, (2) в плазмолемму (в составе окаймленных пузырьков) и (3) в секреторные гранулы (в виде окаймленных пузырьков, утрачивающих в дальнейшем оболочку).
АППАРАТ ВНУТРИКЛЕТОЧНОГО ПЕРЕВАРИВАНИЯ: ЭНДОСОМЫ И ЛИЗОСОМЫ
Аппарат внутриклеточного переваривания представлен системой особых органелл - мембранных пузырьков с кислым содержимым - эндосом (от греч. endo - внутри и soma - тело) и лизосом (от греч. lysis - разрушение и soma - тело), которые обеспечивают катаболические процессы в цитоплазме клетки (рис. 3-11). Функция аппарата внутриклеточного переваривания состоит в регулируемом внутриклеточном расщеплении макромолекул внеклеточного и внутриклеточного происхождения.
Содержание эндосом и лизосом неодинаково в клетках различных типов; оно максимально в тех из них, которые активно осуществляют пиноцитоз и фагоцитоз с последующим перевариванием захваченного материала (в фагоцитах, остеокластах, антиген-представляющих клетках некоторых эпителиоцитах).
Объединение эндосом и лизосом в единую систему основано на наличии в их мембране АТФ-зависимого протонного насоса, вызывающего закисление среды внутри этих органелл. Низкие значения pH активируют ферменты - кислые гидролазы, которые транспортируются особыми гидролазными пузырьками, образующимися в комплексе Гольджи.
Мембрана эндосом и лизосом (толщиной около 6 нм) помимо наличия протонного насоса обладает рядом других важных свойств:
- 52 -
(1)она содержит рецепторы, обусловливающие ее связывание с мембраной гндролазных и транспортных пузырьков, а также фагосом,
(2)обеспечивает свободную диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму, (3) в неповрежденном со-стоянии представляет собой барьер, резистентный к действию литических ферментов и препятствующий их утечке в гиалоплазму.
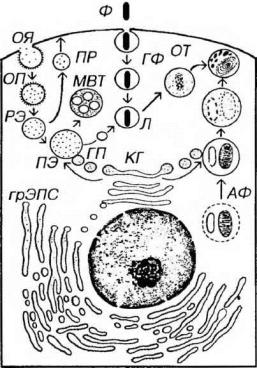
Рис. 3-11. Аппарат внутриклеточного переваривания: эндосомы и лизосомы. КГ
-комплекс Гольджи, ГП - гидролазные пузырьки, ОЯ - окаймленная ямка, ОП - окаймленный пузырек, РЭ - ранняя эндосома, ПР - пузырек рециклирования, ПЭ
-поздняя эндосома, Л - лизосома, ГФ - гетерофагосома, АФ - аутофагосома, ОТ
-остаточное тельце, МВТ - мультивезикулярное тельце.
Эта мембрана стабилизируется гормонами кортикостероидами, а ее повреждение (в результате осмотического воздействия, замораживанияоттаивания, действия ультразвука, высокой температуры, некоторых веществ и др.) приводит к разрушению клетки вследствие самопереваривания литическими ферментами.
- 53 -
Эндосомы
Эндосомы - мембранные пузырьки с постепенно закисляющимся содержимым, которые обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации. В связи с указанными свойствами совокупность эндосом в настоящее время считают не просто механизмом транспорта веществ в клетке (как полагали ранее), а частью системы их переваривания ("внутриклеточного пищеварительного тракта"), в которую входят также лизосомы.
Процесс переноса веществ системой эндосом (по эндоцитозному пути)
может протекать (а) с полным перевариванием макромолекул, (б) с их частичным расщеплением или (в) без изменений по ходу транспорта в лизосому. Способность к перевариванию в эндосомах обеспечивается благодаря тому, что кислые гидролазы вносятся в эндоцитозный путь уже на самых ранних его этапах.
Путь транспорта и деградации веществ в клетке можно описать последовательностью; ранняя (периферическая) эндосома -> поздняя (перинуклеарная) эндосома -> лизосома.
Условия расщепления макромолекул на указанном пути их переноса последовательно становятся все более жесткими. Эндосомы обеспечивают сравнительно мягкий контролируемый прелизосомальный этап переваривания, который необходим и достаточен для легко расщепляемых веществ и комплексов. Наибольшая активность и степень деградации веществ характерна для лизосом, куда переносятся наименее перевариваемые материалы. Благодаря такому устройству клетка располагает органеллами с широким спектром условий расщепления веществ.
Механизм перемещения веществ по эндоцитозному пути остается недостаточно понятным и описывается двумя моделями.
(1)модель челночных пузырьков основана на представлении о переносе поглощенных веществ между стабильными органеллами посредством транспортных пузырьков;
(2)модель созревания предполагает последовательное превращение ("созревание") одной органеллы в другую в пределах указанного пути (компоненты, необходимые для процесса созревания, например, гидролазы, доставляются пузырьками, сливающимися с созревающими эндо-сомами).
-54 -
Ранние (периферические) эндосомы являются мембранными пузырьками на ранних этапах после их отделения от плазмолеммы (но уже после утраты первоначально имевшейся клатриновой оболочки). Они располагаются неподалеку от плазмолеммы в периферических отделах цитоплазмы (см. рис. 3-11). В них в условиях слабокислой среды (pH 6.0) осуществляется ограниченное и регулируемое переваривание макромолекул протеазами, которые были внесены в эндосому, по-видимому, еще на этапе ее формирования. В ранней эндосоме происходит отщепление лигандов от рецепторов с их сортировкой и возможным возвращением последних в специальных пузырьках в плазмолемму для повторного цикла их использования (рециклирования - от англ. recycling). В частности, в эндосоме происходит расщепление комплексов рецептор-гормон (для пептидных гормонов), рецептор-фактор роста, антиген-антитело, а также ограниченный протеолиз (процессинг) антигенов, инактивация или активация ряда молекул. В этой связи раннюю эндосому называют также CVRL (сокр. от англ. Compartment for Uncoupling Receptors and Ligands - компартмент для разделения рецепторов и лигандов).
Поздние (перинуклеарные) эндосомы получили свое название благодаря тому, что они образуются позднее ранних и располагаются в глубоких отделах цитоплазмы вблизи ядра. Они достигают диаметра 600-800 нм и характеризуются сравнительно плотным матриксом. Их отличает от ранних эндосом более кислое содержимое (pH 5.5) и более глубокий уровень переваривания ферментами. В них из ранних эндосом поступают продукты (лиганды), которые должны подвергнуться расщеплению. Большая часть этих продуктов, а также ферменты в дальнейшем будут направлены в лизосому (см. рис. 3-11). Предполагают, однако, что некоторые молекулы могут рециклироваться из поздней эндосомы в раннюю или даже направляться в плазмолемму.
Терминология отдельных компонентов системы эндосом и лизосом еще окончательно не сложилась в связи с активно проводимыми в настоящее время исследованиями в этой области и пересмотром некоторых ранее принятых представлений. Поэтому поздние эндосомы некоторые авторы именуют эндолизосомами, или ранними лизосомами (поскольку они активно участвуют в процессах расщепления веществ). Иногда эндолизосомы выделяют в качестве последнего самостоятельного прелизосомального компонента, входящего в эндоцитозный путь.
Лизосомы ранее традиционно подразделялись на первичные (неактивные) и вторичные (активные). В настоящее время в связи с усложнением представлений о системе эндосом и лизосом использование этих терминов считается более нецелесообразным.
- 55 -
Гидролазные пузырьки (ранее называвшиеся первичными лизосомами) - округлые мембранные органеллы диаметром до 200-400 нм с мелкозернистым плотным матриксом, содержащие литические ферменты в неактивной форме. В большинстве клеток они имеют очень малые размеры (порядка 50 нм), а их надежная идентификация возможна лишь путем демонстрации содержащихся в них ферментов. В фагоцитах они достигают наиболее крупных размеров (до 500 нм). Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки участвуют в транспорте литических ферментов в эндоцитозный путь из сети транс-Гольджи, где они подвергаются окончательным химическим преобразованиям и упаковываются в мембраны.
Литические ферменты гадролазных пузырьков синтезируются и накашиваются в ЭПС, далее переносятся в комплекс Гольджи, где модифицируются и упаковываются в мембранные пузырьки, окруженные клатриновой оболочкой, впоследствии исчезающей. Они содержат олигосахаридные цепочки, имеющие маркер, благодаря которому они направляются не по общему секреторному пути, а сегрегируются в гидролазных пузырьках. В настоящее врет известно около 60 таких ферментов; все они представляют собой кислые гидролазы (гидролитические ферменты с оптимумом рН~5) и включают протеазы, нуклеазы, гликозидазы, липазы, фосфорилазы, фосфат азы и сульфатазы. Ферментный состав гадролазных пузырьков, эндосом и лизосом неодинаков в клетках разных типов; он может различаться даже в отдельных эндосомах и лизосомах одной клетки. Около 20% литических ферментов встроено в мембрану гидролазных пузырьков и временно инактивировано в ней благодаря связи с липидами, примерно 80% находится в матриксе и также инактивировано вследствие отсутствия кислой среды и наличия в их молекуле углеводов. При случайной утечке небольшого количества ферментов из пузырьков отсутствие кислой среды в гиалоплазме защищает ее от разрушения.
Лизосомы (ранее называемые вторичными лизосомами) - органеллы, активно участвующие в завершающих этапах процесса внутриклеточного переваривания захваченных клеткой макромолекул посредством широкого спектра литических ферментов при низких значениях pH (5.0 и ниже). Они формируются с участием поздних эндосом. Диаметр лизосом обычно составляет 0.5-2 мкм, а их форма и структура могут существенно варьировать в зависимости от характера перевариваемого материала. Как и в случае гидролазных пузырьков, они достоверно идентифицируются только на основании выявления в них гидролитических ферментов. Название отдельных видов лизосом основано на наличии в их просвете морфологически распознаваемого материала;
- 56 -
в его отсутствие используется общий термин лизосома. После переваривания содержимого лизосомы образующиеся низкомолекулярные вещества диффундируют через ее мембрану в гиалоплазму.
1)Фаголизосома формируется путем слияния поздней эндосомы или лизосомы с фагосомой, называемой также гетерофагосомой (от греч. heteros - другой, phagein - поедать и soma - тело) - мембранного пузырька, содержащего материал, захваченный клеткой извне и подлежащий внутриклеточному перевариванию; процесс разрушения этого материала называется гетерофагией;
2)Аутофаголизосома образуется при слиянии поздней эндосомы или лизосомы с аутофагосомой (от 1реч. autos - сам, phagein - поедать и soma - тело)
-мембранным пузырьком, содержащим собственные компоненты клетки, подлежащие разрушению. Процесс переваривания этого материала называют аутофагией. Источником мембраны, окружающей клеточные компоненты, служит грЭПС.
3)Мулътивезикулярное тельце (от лат. multi - много и vesicula - пузырек)
представляет собой крупную (диаметром 200-800 нм) сферическую окруженную мембраной вакуоль, содержащую мелкие (40-80 нм) пузырьки, погруженные с светлый или умеренно плотный матрикс. Оно образуются в результате слияния ранних эндосом с поздней, причем мелкие пузырьки формируются, вероятно, путем отпочковывания внутрь от мембраны вакуоли. Матрикс тельца содержит литические фермента и, очевидно, обеспечивает постепенное разрушение внутренних пузырьков.
4)Остаточные тельца - лизосомы, содержащие непереваренный материал, которые могут длительно находиться в цитоплазме или выделять свое содержимое за пределы клетки. Распространенным типом остаточных телец в организме человека являются липофусциновые гранулы - мембранные пузырьки диаметром 0.3-3 мкм, содержащие труднорастворимый коричневый эндогенный пигмент липофусцин. Под электронным микроскопом липофусциновые гранулы представляют собой структуры вариабельной формы, содержащие липидные капли, плотные гранулы и пластинки. В связи с их накоплением в некоторых клетках (нейронах, кардиомиоцитах) при старении, липофусцин рассматривают как "пигмент старения" или "изнашивания".
Секреция лизосомальных ферментов за пределы клетки осуществляется у остеокластов - клеток, разрушающих костную ткань, а также фагоцитов (нейтрофилов и макрофагов) при внеклеточном переваривании различных объектов. Избыточная секреция этих ферментов может приводить к повреждениям окружающих тканей.
- 57 -
Роль гетерофагии в нормальной деятельности клеток и значение ее нарушений. Гетерофагия играет очень важную роль в функции клеток всех тканей и органов. Дефицит тех или иных лизосомальных ферментов (обычно обусловленный наследственными аномалиями) может приводить к развитию ряда заболеваний, вызванных накоплением в клетках непереваренных веществ (чаще всего гликогена, гликолипидов, гликозаминогликанов), которые нарушают их функцию (болезни накопления). При наиболее распространенных заболеваниях, относящихся к этой группе, повреждаются нейроны, макрофага, фибробласты и остеобласты, что клинически проявляется разнообразными по тяжести нарушениями строения и функции скелета, нервной системы, печени, селезенки.
В почке в результате гетерофагии клетки захватывают белки из просвета канальцев и расщепляют их до аминокислот, которые далее возвращаются в кровь. Гетерофагия в клетках щитовидной железы (тироцитах) обеспечивает отщепление йодсодержащих гормонов от белковой матрицы и последующее всасывание их в кровь. Нарушение процесса гетерофагии в указанных клетках вызывает тяжелые расстройства функции этих органов.
Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, в основе деятельности которых лежит поглощение извне и переваривание частиц или веществ. Так, фагоциты (макрофаги и нейтрофильные лейкоциты) захватывают и переваривают микроорганизмы, попадающие в ткани макроорганизма или на их поверхность (например, эпителия слизистых оболочек). При отсутствии или недостаточной активности лизосомальных ферментов, разрушающих микробы (например, при ряде генетически обусловленных нарушений), эти клетки неспособны эффективно осуществлять защитные функции, что приводит к развитию тяжелых хронических воспалительных заболеваний.
Наиболее патогенные микроорганизмы ускользают от повреждающего действия фагоцитов, осуществляя это различным образом. Так, одни (например, возбудитель проказы) обладают устойчивостью к действию лизосомальных ферментов; другие микробы (например, возбудитель туберкулеза) способны подавлять процесс слияния фагосом с ли зосомами, некоторые могут ускользать от разрушения, разрывая мембраны фагосом или лизосом.
Роль аутофагии в нормальной деятельности клеток и значение ее нарушений. Аутофагия обеспечивает постоянное обновление ("омоложение") клеточных структур благодаря перевариванию участков цитоплазмы, митохондрий, скоплений рибосом, фрагментов мембраны (убыль которых компенсируется их новообразованием). Этот процесс обновления в клетке тонко отрегулирован, причем каждый ее компо-
- 58 -
нент имеет определенную продолжительность жизни. Так, в нейронах пожилого человека, которые функционировали на протяжении многих десятилетий, большинство органелл не старше 1 мес. В клетках печени (гепатоцитах) большая часть цитоплазмы разрушается менее, чем за 1 нед. В некоторых случаях аутофагия может служить реакцией клетки на недостаточное питание. Частным случаем аутофагаи является кринофагия (от греч. krinein - отделяю, секретирую) - лизосомальное разрушение избытка невыведенного секрета в железистых клетках.
ПЕРОКСИСОМЫ
Пероксисомы (микротельца) по своему строению сходны с лизосомами. Они представляют собой мембранные сферические или удлиненные пузырьки диаметром 0.05-1.5 мкм с умеренно плотным однородным или мелкозернистым содержимым (матриксом), в котором иногда выявляется более плотная сердцевина (нуклеоид), имеющая кристаллическое строение и состоящая из фибрилл и трубочек. Мелкие пероксисомы (микропероксисомы) диаметром 0.05-0.25 мкм встречаются во всех клетках, крупные (макропероксисомы) диаметром 0.3-1.5 мкм - в гепатоцитах, макрофагах, клетках проксимальных почечных канальцев. Число пероксисом варьирует в клетках разных типов; в гепатоцитах оно составляет в среднем 500, а занимаемый ими относительный объем - около 2% объема клетки. Пероксисомы обновляются каждые 5-6 дней.
Матрикс пероксисом содержит до 15 ферментов, состав которых может варьировать. Наиболее важные из них - пероксндаза, каталаза (на которую приходится до 40% общего белка органеллы), оксида за Б-аминокислот и уратоксидаза. Нуклеоид пероксисомы соответствует области конденсации ферментов.
Образование пероксисом происходит в ЭПС, путем отпочковывания от элементов аЭПС; их ферменты синтезируются частично в грЭПС, частично - в гиалоплазме. По некоторым представлениям, пероксисомы образуются вследствие расщепления ранее существующих, растущих благодаря постоянному поступлению ферментов. Мембрана пероксисомы высокопроницаема для ионов и низкомолекулярных субстратов.
Функции пероксисом. Пероксисомы (наряду с митохондриями) - главный центр утилизации кислорода в клетке. В результате окисления аминокислот, углеводов и других соединений в клетках образуется сильный окислитель - перекись водорода (Н2O2), которая далее благодаря действию каталазы пероксисом распадается с выделением кислорода и воды. Пероксисомы защищают клетку от действия перекиси водорода, оказывающей сильный повреждающий эффект.
- 59 -
Крупные пероксисомы печени и почек играют важную роль в обезвреживании ряда веществ. Например, в них окисляется около 50% поглощенного этилового спирта. Помимо реакций детоксикации, ферменты пероксисом катализируют расщепление жирных кислот, участвуют в ряде катаболических и анаболических реакций, в частности, в обмене аминокислот, оксалата и полиаминов. Некоторые из этих реакций протекают исключительно в пероксисомах, отчего их повреждение может привести к серьезным обменным нарушениям.
В настоящее время открыт новый класс наследственных заболеваний человека, насчитывающий не менее 12 нозологических единиц - пероксисомные болезни, развитие которых обусловлено дефектом активности пероксисом. При этих болезнях поражаются различные органы, часто развиваются тяжелые нарушения нервной системы, вызывающие смерть больных в детском возрасте.
ЭНЕРГЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ: МИТОХОНДРИИ
Митохондрии представляют собой мембранные полуавтономные органеллы, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот.
Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Их размеры составляют 0.2-2 мкм в ширину и 2-10 мкм в длину, а количество в различных клетках варьирует в широких пределах, достигая в наиболее активных 500-1000. В клетках печени (гепатоцитах) их число составляет около 800, а занимаемый ими объем равен примерно 20% объема цитоплазмы. На светооптическом уровне митохондрии выявляются в цитоплазме специальными методами и имеют вид мелких зерен и нитей (что обусловило их название - от греч. mitosнить и chondros - зерно).
В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС).
Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны - кристы (рис. 3-12).
- 60 -