

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfответственных за ПД. Дальнейшая деполяризация вызывает уменьшение амплитуды ВПСП, а при потенциале, близком к нулю, ВПСП не возникает. Еще большая деполяризация приводит к инверсии знака потенциала. Подобная зависимость амплитуды и знака ВПСП от мембранного потенциала, а также наличие равновесного потенциала свидетельствует о том, что ВПСП является результатом ионных токов, протекающих через мембрану по своим электрохимическим градиентам. Формально активация возбуждающего синапса соответствует замыканию ключа в электрической схеме (рис. 6.6), в результате чего включается электродвижущая сила (э.д.с.) равновесного потенциала ВПСП.
Ðèñ. 6.7. Токи отдельных каналов, зарегистрированных методом пэтч-кламп, при аппликации АХ на мембрану мышечной клетки (вверху) и суммарный ток концевой пластинки (внизу). Токи отдельных каналов синхронизированы от момента аппликации АХ. Ток концевой пластинки является результатом сложения токов, протекающих через одиночные каналы.
140
Âдальнейшем было установлено, что рецептор, чувствительный к ацетилхолину, является ионотропным и управляет каналом, проводящим при активации смешанный Na+/Ê+-ток (см. параграф 5.4.1) С использованием метода пэтч-кламп было показано, что при непрерывной аппликации АХ активируемые им ионные каналы открываются и закрываются по принципу «все или ничего». Суммарный ток, складывающийся из токов отдельных каналов, синхронизированных от момента аппликации АХ, по форме и длительности соответствует току концевой пластинки (рис. 6.7).
ВПСП в нейронах обладают такими же свойствами, как ПКП в мышечных клетках. По своему функциональному значению в механизмах межнейронной сигнализации ВПСП является событием, повышающим вероятность генерации ПД и, соответственно, распространения возбуждения по нейронным сетям. Ток, который вызывает смещение потенциала на мембране в сторону деполяризации (ВПСП), называют возбуждающим постсинаптическим током (ВПСТ). С развитием микроэлектродной техники были изучены многие ВПСП (и ВПСТ), возникающие при химической синаптической активации всеми известными медиаторами всех известных рецепторов.
6.2.2.Тормозные постсинаптические потенциалы
Âсинапсах, обеспечивающих синаптическое торможение с активацией ионотропных каналов, лежат те же механизмы, что и в основе работы возбуждающих синапсов с активирующими ионотропными рецепторами. В нейроне регистрируется ВПСП, если в его мембране активируются каналы, пропускающие токи с потенциалом реверсии более положительным, чем порог ПД. Напротив, в нейроне будет регистрироваться тормозный потенциал (отклонение в сторону гиперполяризации), если в его мембране активируются каналы, пропускающие токи с потенциалом реверсии более отрицательным, чем порог ПД.
Гиперполяризация, вызываемая тормозным медиатором в нейроне, называется тормозным постсинаптическим потенциалом (ТПСП). По своему функциональному значению в механизмах межнейронной сигнализации ТПСП является событием, снижающим вероятность генерации ПД и, соответственно, прекращающим (или затрудняющим) распространение возбуждения по нейронным сетям. Ток, который вызывает смещение потенциала на мембране в сторону гиперполяризации (ТПСП), называют тормозным постсинаптическим током (ТПСТ).
Прямое синаптическое торможение вызывается активацией тормозным медиатором ионотропных каналов, проводящих ионы Cl–. Во многих нейронах ионы Cl– имеют равновесный потенциал, близкий к ПП.
141
Первые исследования прямого синаптического торможения были проведены на нервно-мышечном соединении членистоногих, рецепторе растяжения рака и мотонейронах спинного мозга кошки.
Мотонейроны спинного мозга млекопитающих тормозятся тормозными интернейронами.
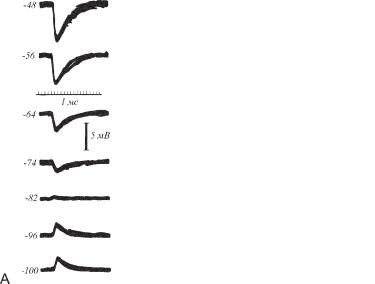
С целью исследования характеристик ТПСП методом фиксации тока изменяли мембранный потенциал мотонейрона и стимулировали его тормозные входы (рис. 6.8 А). При потенциале покоя (-74 мВ) стимуляция тормозных входов вызывает небольшой по амплитуде ТПСП. При деполяризации мембраны (-64 мВ и выше) амплитуда ТПСП монотонно увеличивается. При гиперполяризации клетки до -82 мВ амплитуда ТПСП снижается почти до нуля и при этом меняет полярность (реверсия потенциала). При гиперполяризации мембраны ниже -82 мВ амплитуда реверсированного (положительного по знаку) ТПСП монотонно увеличивается. Таким образом, потенциал реверсии ТПСП в мотонейроне составляет около -80 мВ.
Ðèñ. 6.8. Слева — ТПСП мотонейрона, вызванные электрической стимуляцией тормозных входов, при различном уровне мембранного потенциала, устанавливаемом методом фиксации тока через внутриклеточный электрод. Справа — эквивалентная электрическая схема тормозного синапса. Активация тормозного синапса (замыкание ключа) включает э.д.с. равновесного потенциала ТПСП.
142
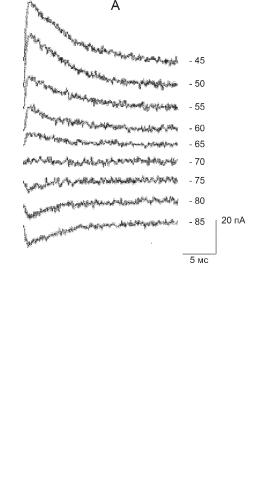
Ðèñ. 6.9. (А) Зависимость ТПСТ мотонейрона, вызываемых стимуляцией тормозных входов, от мембранного потенциала. (Б) Зависимость амплитуды и полярности ТПСТ мотонейрона от мембранного потенциала.
В другом эксперименте посредством метода фиксации потенциала были измерены токи, текущие через мембрану мотонейрона, при стимуляции его тормозных входов (рис. 6.9). При деполяризации мембраны выше -70 мВ регистрировали выходящие токи увеличивающейся
143
амплитуды, отражающие вход анионов Cl– в нейрон. При гиперполяризации мембраны ниже -70 мВ направление тока менялось, а его амплитуда увеличивалась с гиперполяризацией. Таким образом, уровень потенциала -70 мВ является для данного нейрона точкой реверсии Cl–-тока, и, соответственно, ТПСП.
Выявленная зависимость свойств ТПСП от мембранного потенциала и наличие равновесного потенциала свидетельствует в пользу того, что ТПСП является результатом ионного тока, протекающего через мембрану по своему электрохимическому градиенту. Формально тормозный синапс можно представить ключом, который замыкает электрическую цепь, в результате чего включается э.д.с. равновесного потенциала ТПСП (рис. 6.8 Б). Каналы в мембране мотонейронов, ответственные за торможение, пропускают ионы Cl-. Прямым доказательством хлорного механизма ТПСП является тот факт, что введение Cl– в мотонейроны сдвигает хлорный равновесный потенциал и, соответственно, потенциал реверсии ТПСП в положительном направлении.
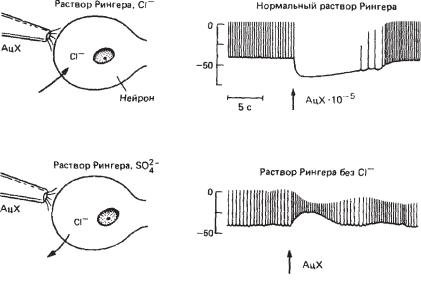
Ðèñ. 6.10. Инверсия ТПСП нейрона улитки, вызванного аппликацией АХ на его мембрану, при смещении равновесного потенциала для Cl– при его замене во внеклеточном растворе на SO42. Вверху — ТПСП в ответ на аппликацию АХ в нормальных условиях, внизу — деполяризация в ответ на аппликацию АХ в среде без Cl–.
144
Аналогичный эксперимент был проведен на нейронах улитки, только вместо введения Cl– в нейрон этот анион удалили из внеклеточной среды, заменив его на SO42-. В естественных условиях при действии АХ на некоторые нейроны улитки (H-клетки) в их мембранах увели- чивается Cl–-проводимость, что вызывает гиперполяризацию (ТПСП) и прекращение импульсной активности (рис. 6.10). При замене во внеклеточной среде Cl– íà SO42– АХ вызывает в Н-нейронах ток противоположного направления из-за того, что равновесный потенциал для Cl– сместился в положительном направлении. В результате вместо ТПСП в нейроне генерируется деполяризация, что приводит к учащению разрядов нейрона.
6.3. Пресинаптическое торможение
Этот дополнительный механизм торможения был открыт в спинном мозге кошки, а также в нервно-мышечном соединении речного рака. Пресинаптическое торможение заключается в уменьшении коли- чества медиатора, высвобождаемого из возбуждающих аксонных терминалей.
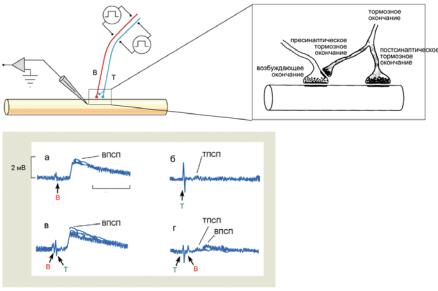
Ðèñ. 6.11. Пресинаптическое торможение в нервно-мышечном синапсе рака. В — возбудительный вход, Т — тормозный вход. Объяснения в тексте.
145
Тормозный нерв в нервно-мышечном соединении рака воздействует не только на мышечное волокно, но также и на аксонную терминаль возбуждающего входа. Стимуляция по отдельности возбуждающего и тормозного входов вызывает, соответственно, ВПСП и малый по амплитуде ТПСП положительной амплитуды (рис. 6.11, а, б). Если возбуждающий вход активируется на 1,5 мс раньше, чем тормозный (рис. 6.11, в), в мышце развивается обычный ВПСП (как на рис. 6.11, а). Если тормозный вход активируется раньше возбудительного (рис. 6.11, г), в мышце регистрируется положительный ТПСП (как на рис. 6.11, б) и следующий за ним ВПСП существенно меньшей, чем на фрагменте а, амплитуды. Пресинаптическое и постсинаптическое окончания тормозного входа выделяют ГАМК, которая увеличивает Cl--проводимость. Предполагается, что при возрастании Cl--проводимости в мембране нервного окончания ослабляется деполяризующий эффект приходящего ПД. В результате происходит снижение амплитуды ПД и количества выделившегося медиатора из возбуждающего окончания.
Было показано, что пресинаптическое торможение во многих синапсах млекопитающих происходит также и в результате ингибирования потенциал-зависимых Са2+-каналов в окончаниях аксонов. Функциональное значение пресинаптического и постсинаптического торможения в ЦНС существенно различаются. Постсинаптическое торможение снижает вероятность генерации ПД, устраняя все возбуждающие влияния. Пресинаптическое торможение избирательно блокирует (или ослабляет) только определенный вход, оставляя возможность нейрону реагировать на другие воздействия. Пресинаптическое торможение осуществляется через аксо-аксональные синапсы, существование которых было показано методом электронной микроскопии в нервно-мышечном соединении рака и в различных областях ЦНС млекопитающих. Пресинаптические эффекты имеют место и на тормозных окончаниях, что было обнаружено в тормозных синапсах рецептора растяжения рака. Кроме того, имеются также данные о существовании пресинаптического возбуждения, которое усиливает деполяризацию пресинаптического окончания и, соответственно, увеличивает количество выделяемого медиатора.
6.4.От чего зависит постсинаптический эффект?
Âреальных условиях нейрон подвергается массированной синаптической бомбардировке со стороны окружающих нейронов. В то же время в самом нейроне и в локальном его окружении происходят трудно контролируемые изменения ионного состава, РН, метаболические процессы, влияющие на состояние нейрона.
146
Эффект медиатора определяется типом рецептора, который управляет каналом, проводящим один или несколько ионов. Знак и амплитуда постсинаптического потенциала зависит от типа иона, который проникает через канал, и его равновесного потенциала. При возникновении проводимости для определенного иона переносимый им ток будет сдвигать мембранный потенциал в сторону равновесного потенциала для этого иона. Равновесный потенциал иона определяется различием концентраций по обе стороны мембраны. Если через активированный канал проходят несколько ионов, то результирующий равновесный потенциал будет аддитивно складываться из равновесных потенциалов для каждого из ионов. Так, равновесный потенциал ПКП в мышечном волокне, каналы которого при активации пропускают ионы Na+ è Ê+, складывается из суммы равновесных потенциалов для Na+ (около +55 мВ) и К+ (около -90 мВ) и в результате оказывается примерно посередине между этими равновесными потенциалами. Значение мембранного потенциала, при котором суммарный ток равен нулю, т.е. входящие токи компенсируется выходящими, называют потенциалом реверсии тока.
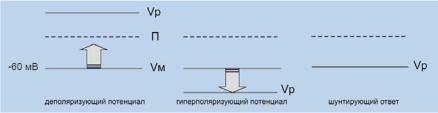
Ðèñ. 6.12. Зависимость эффекта тока от его равновесного потенциала. Vр — потенциал реверсии, Vм — потенциал мембраны, П — порог генерации ПД. Объяснения в тексте.
Если синаптически активируемые токи приближают мембранный потенциал к пороговому уровню ПД (деполяризация), то этот потенциал является возбуждающим (рис. 6.12, слева). Если токи отдаляют мембранный потенциал от порогового уровня ПД (гиперполяризация), то этот потенциал является тормозным (рис. 6.12, в середине). Тормозный ответ не обязательно должен быть гиперполяризующим. Например, часто синаптическое торможение заключается в увеличении К+-проводимости. Равновесный потенциал для К+ примерно равен ПП, и при активации К+- проводимости обычно не происходит изменений величины мембранного потенциала (рис. 6.12, справа). Однако этот эффект действительно является тормозным и значительно ослабляет возбуждающие воздействия на нейрон. В этом заключается шунтирующее действие К+-проводимости. При
147
этом не регистрируются какие-либо выходящие токи, но открытие К+ каналов приводит к существенному падению сопротивления мембраны, и, в соответствии с законом Ома, требуются существенно большие входящие токи, чтобы сместить потенциал мембраны к порогу генерации ПД. Эффективность синаптического воздействия может меняться и при изменениях равновесных потенциалов для ионов. В свою очередь равновесные потенциалы зависят от градиента концентрации для ионов по обе стороны мембраны, которые могут варьировать по многим причинам.
6.5. Потенциалы непрямой синаптической передачи
Синаптические эффекты, опосредованные метаботропными рецепторами, отличаются замедленной динамикой развития потенциалов, поскольку метаболические процессы включают сложные многоэтапные внутриклеточные каскады, требующие большего времени (параграф 5.3).
Многие медиаторы активируют метаботропные рецепторы, которые увеличивают К+-проводимость в мембранах нейронов. Стимуляция тормозных интернейронов в коре мозга через возбудительные афференты вызывает выделение ГАМК в синапсе на пирамидном нейроне (рис. 6.13, А). Сразу после стимуляции в пирамидном нейроне регистрируется ранний ВПСП, поскольку коллатераль возбудительного входа активирует и пирамидный нейрон (рис. 6.13, Б). Тормозный эффект ГАМК в пирамидном нейроне представлен ранним и поздним ТПСП, которые обеспечиваются, соответственно, рецепторами ГАМКÀ è ÃÀÌÊÂ. Ионотропный ГАМКÀ-рецептор вызывает ранний и непродолжительный ТПСП, регистрируемый сразу после ВПСП, который успевает развиться из-за дополнительной синаптической задержки на интернейроне. Однако после раннего ТПСП мембрана не возвращается к ПП из-за генерации позднего ТПСП. Локальное применение антагониста ГАМКÂ-рецепторов факлофена устраняет поздний ТПСП, подтверждая К+-природу позднего ТПСП, обусловленного активацией ГАМКÂ-рецептора. Устранение факлофена приводит к восстановлению позднего ТПСП.
6.6. Быстрые и медленные постсинаптические потенциалы
Разные медиаторы, воздействуя на разные рецепторы, вызывают в нейронах и в других эффекторах разнообразные ПСП, выполняющие множество функций в механизмах нейронной интеграции. Эти потенциалы отличаются по эффекту (возбудительные и тормозные) и временной динамике (быстрые и медленные). Ниже приведена краткая характеристика некоторых изученных постсинаптических потенциалов.
148
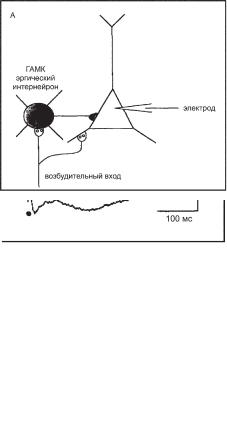
Ðèñ. 6.13. Торможение пирамидного нейрона через ионотропные и метаботропные рецепторы ГАМК. (А) Конфигурация возбудительных (светлые) и тормозных (темные) синапсов на тормозном и пирамидном нейронах. (Б) Сразу после стимуляции в пирамидном нейроне регистрируется ранний ВПСП, поскольку коллатераль возбудительного входа активирует и пирамидный нейрон (контроль). Активация ГАМКÀ-рецепторов вызывает увеличение Cl--тока, который обеспечивает ранний быстрый ТПСП. Активация ГАМКÂ-рецепторов приводит к увеличению К+-тока (через метаботропные К+-каналы), который вызывает задержанный продолжительный ТПСП. Антагонист ГАМКÂ-рецепторов факлофен вызывает угнетение задержанного ТПСП (применение факлофена). При удалении факлофена задержанный ТПСП восстанавливается (удаление факлофена).
149