

Биосинтез полипептидной цепи
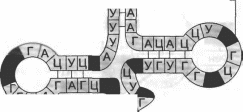
В полипептидной цепи происходят расшифровка информации, закодированной с помощью генетического кода, и построение на матрице мРНК полипептидной цепи определенного белка. В этом процессе участвуют еще два вида РНК — рибосомальная (рРНК) и транспортная (тРНК). Для обоих видов РНК в геноме имеются многочисленные гены, на матрице которых эти РНК синтезируются. Принципиально транскрипция для рРНК и тРНК ничем не отличается от только что описанной транскрипции мРНК. В то же время рРНК и тРНК являются конечными продуктами. Таким образом, в геноме, кроме генов, контролирующих синтез всех белков организма, существуют гены, кодирующие тРНК, рРНК и ряд других типов РНК; тРНК обеспечивает связь между кодонами мРНК и аминокислотами будущей полипептидной цепи. Для каждой из 20 аминокислот существует не менее одной тРНК. Различные тРНК отличаются по нуклеотидной последовательности и по содержанию некоторых редких нуклеотидов, которые возникают с помощью особой по- сттранскрипционной модификации молекулы тРНК. Все виды тРНК содержат около 80 нуклеотидов и на двухмерном изображении имеют форму клеверного листа. К З’-концу тРНК ковалентно присоединяется ее аминокислота. Внутренняя петля (рис. 3.8) содержит 7 неспаренных оснований и
'Структуры, которые осуществляют реакции сплайсинга для получения зрелой мРНК, называют сплайсеосомами. Эти структуры представлены двумя типами молекул — малыми ядерными РНК и малыми ядер- ными рибонуклеопротеинами (мяРНП). которые образуются из малых ядерных РНК и белков. Известно 5 типов мяРНП, которые необходимы для нормального осуществления реакций сплайсинга.
Аминокислота
З'-конец
У
5'-конец
У
г-
Г-У
Модифицированные
нуклеотиды
Петля 1
Петля 3
 .
.
|
|
|
ц |
|
f] |
ц |
- |
Г |
А |
|
У |
|
|
|
Петля
2
Антикодон
Рис. 3.8. Молекула транспортной РНК
Вторичная структура тРНК имеет вид клеверного листа Внутренняя петля (петля 2) содержит 7 неспаренных оснований и антикодон Аминокислота присоединяется к З’-концу тРНК
антикодон — тройку нуклеотидов, комплементарных кодону мРНК. Комплементарность антикодона тРНК кодону мРНК является тем специфическим механизмом, который обеспечивает строгую реализацию последовательности кодонов мРНК и соответственно гена, в последовательность аминокислот кодируемой им полипептидной цепи. Связывание аминокислоты с соответствующей тРНК обеспечивается особым ферментом, который называют аминоацил-тРНК-синте- тазой. Для каждой аминокислоты имеется особая форма этого фермента. Фермент узнает свою аминокислоту и свою
ДНК
АААЦТЦЦАЦТ-ГЦТ+Ц УУУГАГГУГААГААГ
мРНК
Ядерная
мембрана
— г
У*- ь
i Направление движения рибосомы
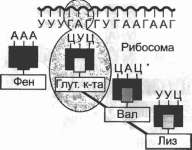
Рис. 3.9. Синтез полипептиднои цепи на рибосоме
Показаны также транскрипция мРНК и ее перенос через ядерную мембрану в цитоплазму клетки.
Формирующаяся полипептидная цепь
тРНК и ковалентно связывает их между собой. В результате образуется комплекс, который называют аминоацил-тРНК. Аминоацил-тРНК-синтетаза обеспечивает также контроль за правильностью соединения тРНК со своей аминокислотой и способна исправить ошибку в случае ее возникновения.
Образование полипептидной цепи из последовательно доставляемых к мРНК тРНК с соответствующими аминокислотами происходит на рибосомах (рис 3.9). Рибосомы представляют собой нуклеопротеидные структуры, в которые входят три вида рРНК и более 50 специфических рибосомных белков. Рибосомы состоят из малой и большой субъединиц Инициация синтеза полипептидной цепи начинается с присоединения малой субъединицы рибосомы к центру связывания на мРНК и всегда происходит при участии метионино- вой тРНК особого типа, которая связывается с метионино- вым кодоном АУГ и прикрепляется к так называемому Р-уча- стку большой субъединицы рибосомы. Следующий кодон
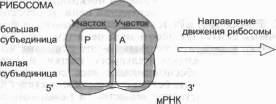
Рис.
3.10. Синтез
полипептидной цепи на рибосоме.
Детализованная
схема присоединения к растущей
полипептидной цепи новой аминокислоты
и участие в этом процессе участков А
и Р
большой субъединицы рибосомы.
Объяснение в тексте.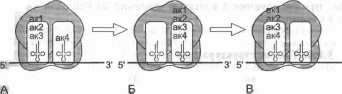
мРНК, расположенный вслед за АУГ-инициирующим кодоном, попадает в A-участок большой субъединицы рибосомы, где он «подставляется» для взаимодействия с амино- ацил-тРНК, имеющей соответствующий антикодон. После того как подходящая тРНК связалась с кодоном мРНК, находящимся в A-участке, происходит образование пептидной связи с помощью пептидилтрансферазы, входящей в состав большой субъединицы рибосомы, и аминоацил-тРНК превращается в пептидил-тРНК. Это заставляет рибосому продвинуться на один кодон, переместить образованную пепти- дил-тРНК в P-участок и* освободить A-участок, который занимает следующий по порядку кодон мРНК, готовый к соединению с аминоацил-тРНК, имеющей подходящий антикодон (рис. 3.10). Происходит рост полипептидной цепи за счет многократного повторения описанного процесса. Рибосома движется вдоль мРНК, высвобождая ее инициирующий участок. На инициирующем участке происходит сборка следующего активного рибосомного комплекса и начинается синтез новой полипептидной цепи. Таким образом к одной молекуле мРНК может присоединиться несколько активных рибосом с образованием полисомы. Синтез полипептида продолжается до тех пор, пока в A-участке не окажется один из трех стоп-кодонов. Стоп-кодон распознается специализированным белком терминации, который прекращает синтез и способствует отделению полипептидной цепи от рибосомы и от мРНК. Рибосома и мРНК также разъединяются и готовы начать новый синтез полипептидной цепи (см. рис. 3.9). Остается только напомнить, что белки — это основные молекулы, обеспечивающие жизнедеятельность клетки и организма. Они и ферменты, обеспечивающие весь сложнейший обмен веществ, и структурные белки, составляющие скелет клетки и образующие межклеточное вещество, и белки-транспортеры многих веществ в организме, как, например, гемоглобин, транспортирующий кислород и белки-каналы, обеспечивающие проникновение в клетку и удаление из нее разнообразных соединений.