

2. Паразитизм: происхождение и распространение. Виды паразитов. Влияние паразитов на организм хозяина.
Паразитизм — исторически сложившаяся ассоциация генетически разнородных организмов, основанная на иммунобиологических взаимоотношениях, пищевых связях и взаимообмене, при котором один (паразит) использует другого (хозяина) в качестве среды обитания и источника питания, причиняя ему вред.
Крупнейший гельминтолог Р. С. Шульц, характеризуя сущность паразитизма, в основу первого критерия взял место обитания или пространственные отношения, второго — питание (метаболическая связь) и третьего — патогенное действие на организм хозяина, вытекающее из первых двух.
Первая попытка охарактеризовать паразитизм принадлежит И. И. Мечникову (1874). Он говорил, что настоящими паразитами называются такие существа, которые живут и питаются за счет других организмов. Лейкарт подтвердил (1879) высказывания И. И. Мечникова.
Если паразитизм и его разнообразие у животных и человека изучены довольно подробно, то его происхождение и становление до сих пор остаются во многом малоизвестными. Это связано с тем, что эволюция животных — хозяев паразитов шла не сплошной линией через геологические эры, а во многом фрагментарно. Тем не менее понимания происхождения паразитизма необходимо изучить рию эволюции животных - дефинитивных и промежуточных
Ученые доказали незаменимость палеонтологических методов для изучения возникновения животного мира на Земле. Так, в период кембрия (580-570 млн лет назад) появляются предки пауков, скорпионов, клещей и других ™исто-ногих и предки моллюсков, раков, иглокожих и т. п Начиная от ордовика и до перми (от 500 до 285-230 млн лет назад) бурно развивались как беспозвоночные, так и позвоночные животные. Гельминты, как и другие паразиты, встречались уже в первобыт и стегоцефалов, просуществовавших многие милл. лет. То же самое можно предположить и о рептилиях 14 отрядов которые погибли 65 млн лет назад. Однако вместе с «хозяевами» вымерли и их специфические паразиты. В то же время представители отдельных видов (кистеперая рыба) и отрядов (гатте-рти черепахи, змеи и крокодилы), дошедшие до нынешних времен, Логически должны иметь и «своих» давних паразитов с последующим пополнением видов. Организмы, паразитирующие в домашних и промысловых животных, могли появиться у них по мере их становления (за период 70— 0,25—0,01 млн лет назад).
Эволюция природы шла по принципу увеличения разнообразия представГелей растительного и животного мира, совершенствована их взаимоотношений. Со временем возникали более сложные взаимоотношения животных разных видов. Более простые взаимоотношения (нахлебничество и др.) со временем могли перерасти в одну из форм паразитизма.
Об этом свидетельствуют следующие положения: адаптация к различным типам и способам питания представителей классов , эндо-и эктопаразитов; монофункциональные изменения , в процессе эволюции как наружных, так и внутренних органов паразитов, слож ный цикл развития со сменой хозяев и среды при постоянстве генома особенно хорошо прослеживаемого у эндопаразитов; строгая п^риспособленность (специфичность) паразитов к обитанию на животных определенных видов.
Можно легко проследить генезис эктопаразитов, возникших через разновидности симбиоза, на примере усоногих рачков Cirripedia. Они перешли к эктопаразитизму черезУсинойкию, но их свободноживушие формы ведут сидячий обЬаз жизни Пухоеды тоже перешли к эктопаразитизму от свободноживущих форм в гнездах птиц. Переход к облигатному эктопаразитизму, через хищничество просматривается на примере пиявок (рыбья пиявка Piscicola geometra).
Ученые приводят примеры, когда эктопаразиты в процессе филогении становятся эндопаразитами. Например, паразит мочевого пузыря лягушки некогда обитал на жабрах бесхвостых амфибий. Примером перехода хищничества в паразитизм могут служить клопы. Среди них есть типичные паразиты (семейство Cimecidae) и хищники (Reduvius personatus).
Возникновение эндопаразитизма проходит поэтапно: регулирование кислородного обмена в анаэробных условиях; снижение затрат на подвижность и обмен; обеспечение незаменимым строительным материалом интенсивного биосинтеза, связанного с быстрым ростом и крайне высокой активностью репродуктивной системы; преодоление защитных барьеров и систем организма хозяина и т. д.
Согласно современным представлениям турбеллярии произошли от древних кишечнополостных, а так как трематоды и цестоды произошли от турбеллярии, то древние кишечнополостные — далекие предки плоских гельминтов
В целом у Scolecida (надтип плоских гельминтов) паразитизм распространен неравномерно (табл. 1). Некоторые классы полностью представлены паразитическими организмами, включая экто- и эндопаразитов, число которых может быть различным. Мезозой — эндопаразиты беспозвоночных — прошли путь глубокой морфо-функциональной регрессии и представлены небольшим числом видов. Моногенеи — в основном эктопаризиты рыб и амфибий, гиродактилиды — эндопаразиты рыб (акуловых и химер) тоже представлены небольшим количеством, но, безусловно, относятся к гельминтам одной из древних групп.
1. Численность видов паразитических платод и скребней
(по Ф. Ф. Сопрунову)
|
Класс |
Общее ЧИСЛО |
Эктопаразиты |
Эндопаразиты |
Развитие |
Окончательный хозяин |
|
Turbellaria |
1600 |
80 |
_ |
Прямое |
Беспозвоночные |
|
Temnocephala |
50 |
20 |
— |
» |
» |
|
Trematoda |
Более |
— |
Более |
Сложное |
Позвоночные |
|
|
3000 |
|
3000 |
|
|
|
Mesozoa |
60 |
— |
60 |
Прямое |
Беспо звоночные |
|
Monogenea |
1500 |
1500 |
— _ |
» |
Позвоночные |
|
Gyrocotylida |
20 |
— |
20 |
» |
» |
|
Cestoda |
Более |
_ |
Более |
Сложное |
» |
|
|
3000 |
|
3000 |
|
|
|
Acanthocephala |
300 |
— |
300 |
» |
» |
Все представители 300 видов известных колючеголовых ведут эндопаразитический образ жизни. Что же касается видов класса нематода, то среди них встречаются хищники, свободноживущие и эндопаразиты животных и человека как с прямым, так и со сложным путем развития. Представители класса мастигофора (жгутиконосцы) по образу жизни и морфологии весьма разнообразны.
Основной признак — это наличие жгутиков в качестве органов движения. По характеру питания и обмена веществ они могут быть животными (зоомастигины) и растительными (фитомастигины) жгутиконосцами. Животные жгутиконосцы питаются мелкими организмами, получая органические вещества в готовом виде, растительные — создают их сами с помощью солнечной энергии. Но есть и такие жгутиконосцы (эвглена зеленая), которые питаются и как растения, и как животные (в зависимости от освещенности), — сапрофиты. Зоомастигины являются объектом изучения ветеринарной и медицинской паразитологии (трипаносомы, трихомонады у животных, лямблии у человека и т. п.).
ПАРАЗИТО-ХОЗЯИННЫЕ ОТНОШЕНИЯ
Любой паразит в организме хозяина — не индифферентное физическое тело, а биологически активный агент, входящий в тесный контакт с ним на физико-химическом и иммунобиологическом уровнях. Этот процесс особенно остро проявляется в системе паразит — хозяин у эндопаразитов. Сложность отношений обусловливается рядом признаков, в том числе строением внешних покровов трематод, цестод и нематод. Так, у паразитических червей первых двух групп внешние покровы представляют собой цитоплазматический тегумент (Rothm частей. Тегумент в отличии кутикулы нематод (особенно у цестод) — активный орган, выполняющий функции секреции, пищеварения и всасывания (рис. 4, 5).
У всех цестод и личинок трематод (у спороцист и частично у редий) на цитоплазматической мембране расположены микроворсинки длиной 600—1500 мкм и шириной 100—400 мкм. Такая структура поверхности тела во много раз увеличивает площадь соприкосновения паразитов с пищевым субстратом и поверхностью окружающей среды, что создает оптимальные условия для усвоения пищи и пристеночного пищеварения (у цестод). У эндопаразитов — цестод и частично трематод — на теле обнаружены все жизненно важные окислительно-восстановительные и пищеварительные ферменты. Эти паразиты способны использовать ферменты хозяина так, что молекула энзимов адсорбируется на поверхности их тела активным концом к окружающей среде. Вследствие этого субстраты организма —химус (у цестод), слизь, тканевая жидкость — при контакте с паразитом им перевариваются и всасываются. Так, неспецифичная щелочная фосфатаза выделена с поверхности тегумента цестод и трематод. Это подтверждает факт активного переноса некоторых питательных веществ через покровы тела.
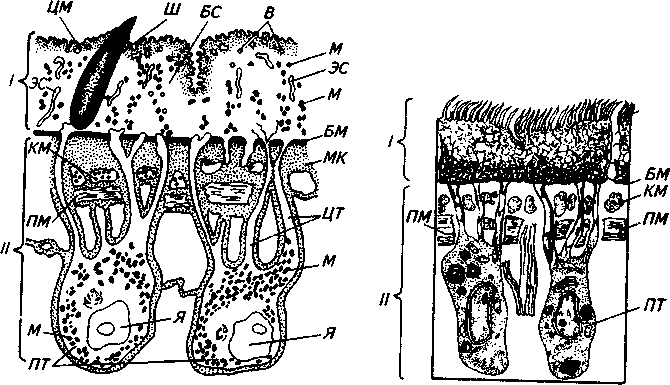
Рис. 4. Строение внешнего покрова (тегумента) трематод:
1— наружная часть: ЦМ — цитоплазматическая мембрана; Ш— шипик; БС—безъядерный синцитий; В — вакуоли; М — митохондрии; ЭС—эндо-плазматическая сеть; БМ — базальная мембрана; // — внутренняя часть: КМ — кольцевые мышцы; ПМ — продольные мышцы; МК—межклеточное вещество; ЦТ — цитоплазматические тяжи; Я — ядра; ПТ— погруженная часть тегумента
Известно, что у цестод отсутствует пищеварительная система в обычном понятии. Функцию пищеварения у них полностью выполняет тегумент. На микроворсинках адсорбируются эстеразы, протеиназы, пептидазы, холин-эстеразы, щелочная фосфатаза и другие ферменты, способные расщеплять пищевые субстраты с последующей транспортировкой продуктов распада в организм паразита. Это доказывает, что тегумент у плоских гельминтов (особенно у цестод) — активно действующий орган и, следовательно, знание функции этого органа поможет в изучении механизма действия антгель-минтиков, их рациональном подборе и использовании.
Холинэстераза была обнаружена у дикроцелиумов и фас-циол в стенках половых органов и мышечных волокнах, проходящих во внутренней части тегумента. Установлено, что у дикроцелиумов хлорофос проникает в организм через внешние покровы, разрушая холинэстеразу и оказывая на трематод губительное действие.
Внешний покров нематод — кожно-мышечный мешок (кутикула). Производное гиподермы, он является многослойным покровом тела и основным барьером для проникновения
различных субстанций, т. е. обладает свойством полупроницаемости (рис. 6).
В стенке кишечника трематод и нематод обнаружены ферменты, способные переваривать соответствующую пищу.
Паразитические простейшие питаются всей поверхностью тела или захватывают частицы пищи ротовым аппаратом (инфузории) или жгутиковым карманом (трипаносомы).
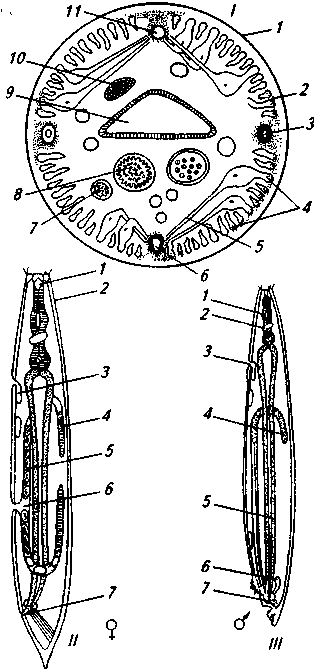
Рис. 6. Строение нематод (по Th. Hiepe):
I— поперечный разрез: 1 — кутикула; 2 — подкутикулярный слой; 3 — экскреторный канал; 4, 5 — мышечные клетки; 6, 11 — вентральный и дорсальный нервные стволы; 7 — яйцевод; 8 — матка, 9 — кишсмник; 10 — яичник; II — самка: / — ротовая капсула; 2 — кутикула; 3 — экскреторная железа; 4 — яичник; 5 —• матка; 6 — вагина; 7 — анальное отверстие; Ш — самец: 1 — пищевод; 2— нервный узел; 3 — экскреторный канал; 4 — семенник; 5 — кишечник; 6 — капсула; 7— клоака
Насекомые и паукообразные имеют развитый ротовой аппарат колющего, сосущего или грызущего типа. У многих из них сильно развиты слюнные железы, при помощи секрета свертываемость крови резко снижается. Если простейшие многих видов обитают в организме хозяина в эпителиальных клетках, эндотелии, мышечных клетках и элементах крови, то демодексы и саркоптесы (хотя они эктопаразиты) паразитируют в толще кожи и ее отдельных элементах.
В целом же паразитические организмы питаются самыми различными по происхождению и составу производными своих хозяев. Например, аскариды человека и свиней, паразитируя в полости тонкого кишечника, питаются его слизистой и химусом. Но их нередко обнаруживают и в желчных протоках печени. Проблема питания здесь остается еще не совсем выясненной.
Немалый интерес во взаимоотношениях паразит — хозяин представляет то, что половые гормоны хозяев-самцов заметно сокращают репродукцию яиц и личинок кишечных нематод, гормоны же хозяев-самок оказывают противоположное действие.
Среди многих проблем взаимоотношений паразит — хозяин есть и такие, которые всегда вызывают живой интерес. Почему, например, в желудке собаки, где перевариваются даже массивные кости, не перевариваются нежные личинки эхинококкуса, ценуруса и других паразитов? Известны отдельные факты, что паразиты вырабатывают биологически активные вещества (ингибиторы), предохраняющие их от переваривания и гибели в организме специфического хозяина. Доказательством служит то, что мертвые мониезии, помещенные в химус из тощей кишки ягнят, при температуре 37—38 °С под действием трипсина и химотрипсина вскоре распадаются на мелкие фрагменты, для живых же цестод подобная среда в естественных условиях служит местом обитания и развития.
Определенную защитную роль приписывают веществам типа муцина, покрывающего поверхность тела трематод и цестод. В ростковой зоне (шейка) цестод и теле личинок изобилуют известковые тельца, которые нейтрализуют избытки кислот и других метаболитов. Значительна роль нативных белков, находящихся у живых гельминтов в биологически активном состоянии. Нативные альбумины, глобулины и другие белки мало подвержены воздействию трипсина, хотя после денатурации различными агентами легко гид-ролизуются.
Не менее интересны отношения между паразитами и хозяевами на уровне органов и тканей хозяина при проникновении в его организм личинок гельминтов. Например, реакция овцы проявляется образованием соединительнотканной капсулы вокруг личинок эхи-нококкуса, свиней — вокруг трихинеллы, крупного рогатого скота — вокруг цистицеркусов. Казалось бы, активная реакция клеток
хозяина изолирует паразита, разрушающего органы. Однако с момента инкапсуляции личинки могут быть в безопасности несколько месяцев и даже лет. Для осуществления своей главной функции — заражения дефинитивных хозяев.
Рассматривая систему паразит - хозяин, необходимо обратить внимание еще на такое понятие, как специфичность и неспецифичность паразитических opfанизмов по отношению к различным хозяевам. У каждого из патеров существуют исторически обусловленные анатомоморфологи46™' физико-химические и биологические приспособительной реакции. Так, у собак паразитируют гельминты: унцинария, анкилостома. Дипилидиум, токсокара парнокопытных гемонхус, нематодирус, диктиокаулюс, непарнокопытных - делафондия; альфортия, оксиурис. Чем генеалогически дальше животные друг одруга тем ярче выражена эта разница. Однако встречаются гельминты-космополиты (трихинеллы), паразитирующие у животных разных видов.
- Принято считать, что 'высокая специфичность - это свидетельство давности отношений ЯаРазит - хозяин и отражение филогении как паразита, так и хозяина не менее специфичность нужно рассматривать как некое Динамичное явление. Например, Фасциола обыкновенная (трематода) бьша заве„зена в Австралию совсем недавно, и из-за отсутст*ия ее европейского промежуточного хозяина (малый прудовик) о*а приспособилась к паразитированию на местном моллюске другой виДа (Lymnaea tomentosa).
Для установления систры паразит - хозяин требуется наличие некоторых условий: хозя**н и паразит должны вступить в контакт друг с другом, это завис^ от Условии внешней среды, поведения сочленов цепи и т. д.; хозяин должен обеспечить все условия для жизнедеятельности паразита (место обитания, питание и т. nj паразит должен быть усто^чив ко многим противодействующим ему реакциям организма хозяйна.
Чем лучше эти условий в отношениях между хозяином и паразитом выполняются, тем хо^ин и паразит специфичнее друг для друга. Они - неотъемлемая часть становления паразитических отношений, выбора хозяина и ^го закрепления.
Примечательно то, что все известные науке стороныпаразито-хозяинных отношений протекают на Фоне общего Физиологического состояния оргаюРма хозяев- Поэтому степень остроты отношений двух видов животного мира и последствии реакции хозяина на паразита зависит от породы, возраста, типа и полноценности кормления животных, усл^™ содержания и т. д. Паразиты же находясь в организме хозя^на (ФеДа пеРвого порядка) зависимы и от условий окружающей следы (cPe«a ВТОРОГО порядка).
Приспосабливаясь к жизни в разных средах обитания, паразитические организмы развивались по пути регресса или прогресса.
Потеря крыльев у блох, вшей и клопов — явление регресса. Этого нельзя сказать о трематодах и цестодах, у которых могут быть потеряны органы движения, но гипертрофировались органы фиксации — присоски, крючки на хоботке или теле. У инфузорий рода Opalina, обитающих в заднем отделе кишечника лягушек, полностью исчезло ротовое отверстие, у свободноживущих же форм оно есть.
Не следует сводить изменения паразитов только к появлению или исчезновению каких-либо органов. Например, у трипаносом, паразитирующих в плазме крови верблюдов, постоянно меняется структура внешней оболочки под влиянием среды хозяина, что способствует возникновению у паразитов оптимальных приспособительных иммунологических реакций.
Прогрессивным признаком, возникшим в процессе развития эндопаразитизма у многих паразитов, можно считать гермафродитизм, встречающийся у представителей классов трематод и цестод. Не менее ярок признак прогресса — хорошо выраженная репродуктивная способность паразитов и многохозяинность (промежуточные, дополнительные, резервуарные, дефинитивные). Так, фас-циола ежесуточно продуцирует 2400 яиц. Далее в результате бесполого размножения (партеногения) в теле промежуточного хозяина-моллюска из каждого яйца образуется около 100 церкариев. Следовательно, каждая фасциола в идеале может дать в сутки более 240 000 личинок. У бычьего цепня в зрелом членике находится более 170 тыс. яиц, свиная аскарида способна дать ежесуточно до 200 000 яиц.
ВОЗДЕЙСТВИЕ ПАРАЗИТОВ НА ОРГАНИЗМ ХОЗЯИНА
Весь комплекс патологических влияний паразита на организм хозяина зависит от многих факторов: вида паразита, его вирулентности, численности, места обитания, биологии развития и_физиоло-гического состояния хозяина. Одно только понятие «физиологическое состояние хозяина» включает ряд факторов, от которых могут зависеть развитие и болезнетворное влияние паразита на организм животного, например: иммунный статус организма, возраст, тип кормления и содержания.
Во взаимоотношениях паразит — хозяин огромное значение имеет и вирулентность (степень болезнетворности) данного паразита. Она зависит от инфекционных свойств паразита и от восприимчивости заражаемого организма хозяина. Вирулентность паразитов может усиливаться при повышении температуры. Например, адо-лескарии фасциол, выращенные при 22—23 "С, вызывали острый фасциолез у кроликов; выращенные при 15—17 °С обусловили только хроническое течение болезни.
Болезнетворное воздействие на организм животного, вызываемое паразитом при заражении, можно разделить на несколько групп: механическое, аллергическое, токсическое, трофическое и инокуляторное.
Механическое воздействие паразита на организм хозяина обусловлено местом его обитания и биологией развития. Ясно, что локализация гельминта в полости кишечника менее ощутима, чем в печени или головном мозге. Кроме того, следует учитывать миграцию личинок (у аскарид) по аскаридному или неаскаридному типу. Механическое влияние клещей при паразитировании на коже крупного рогатого скота менее выражено, чем при миграции личинок подкожных оводов в толще мускулатуры и коже зараженных животных. Локализация крупных пузырей эхинококкусов в паренхиматозных органах животных, ценуросом в головном мозге овцы, диоктофим в почечной лоханке собаки, дирофилярий в желудочках сердца и предсердии плотоядных обусловливает атрофию не только отдельных частей, но и всего органа.
Вухерерии (круглые гельминты длиной от 4 до 10 см) локализуются в лимфатических сосудах и узлах человека, что затрудняет нормальную циркуляцию лимфы и способствует разрастанию соединительной ткани, заканчивающемуся слоновостью конечностей, груди и мошонки. Некоторые гельминты (мониезии, аскариды) закупоривают просвет кишечника, при этом нарушается целостность слизистой, происходит атрофия эпителиальных клеток, брун-неровых желез и т. д. Простейшие, паразитируя в эритроцитах или эпителиальных клетках, значительно разрушают их.
Следует отметить, что механические изменения органов и тканей, как правило, приводят к нарушениям их многочисленных функций. Следовательно, этот процесс нужно рассматривать как морфо-функциональное воздействие паразитов.
Аллергическое воздействие паразитов заключается в том, что в процессе жизнедеятельности они выделяют продукты метаболизма, секреции и экскреции, которые в первую очередь обладают свойствами аллергенов. Аллергены соматического происхождения выделяются в период линьки личинок и их гибели в органах и тканях хозяина, при дестробиляции цестод в период элиминации из организма. Аллергены гельминтов представляют собой сложные соединения — полипептиды, протеины, полисахариды и гликолипиды. Под их влиянием в организме зараженных животных возникает аллергическая реакция (эозинофилия), вырабатывается иммунитет различной степени напряженности.
Токсическое воздействие паразитических организмов в целом изучено слабо. До сих пор никому не удалось выделить у гельминтов токсины. Тем не менее при хроническом течении гельминто-зов в результате нарушения обмена веществ у больных животных нередко отмечают токсикоз. У больных животных ухудшается общее состояние, снижается аппетит, расстраивается функция желудочно-кишечного канала, уменьшается количество эритроцитов, содержание гемоглобина.
Показателями токсикозов также считают уменьшение (в зависимости от времени) содержания в сыворотке крови холинэстеразы, увеличение количества патологически светящихся лейкоцитов. Предполагаемые токсины гельминтов (различные субстраты) способны оказывать цитопатическое влияние на искусственно выращиваемые клетки (перевиваемые клетки амниона человека), раковые клетки Нр-2, первично трипсинизированные клетки фибробластов эмбриона человека и куриные фибробласты.
При массовом нападении мошек у крупного рогатого скота и лошадей под действием гемолитического яда развивается симулио-токсикоз. У саркоцист (простейшие) выделен токсин саркоцистин, вызывающий некроз тканей у многих лабораторных животных. Местные проявления токсического воздействия гельминтов выражаются в дистрофических и некротических изменениях тканей в местах локализации паразитов. Так, при эуритремозе овец в поджелудочной железе можно наблюдать некроз стенки протоков с полным сглаживанием всех структурных элементов. Дегенерация саркоплазмы с потерей исчерченности и глыбчатым распадом отмечается при трихинеллезе.
Трофическое воздействие — неотъемлемое свойство паразита. Если бы известные нам паразитические организмы употребляли ненужные хозяину субстраты типа экскретов или непереваренную пищу, то их следовало бы считать какой-либо разновидностью комменсализма. Способы питания и пища, которую паразиты потребляют из организма хозяина, многообразны и до конца не изучены.
В целом цестоды, как уже было сказано ранее, питаются всей поверхностью благодаря структуре и приспособленности гельминтов использовать пищеварительные ферменты хозяина. У трематод имеется развитая пищеварительная система, и в определенной степени они способны переваривать при помощи специфических ферментов самые разные виды субстратов: кровь, тканевые соки, слизь, эпителий и т. д.
Паразиты с большой биомассой, безусловно, используют значительную часть пищи из организма хозяина. Они потребляют не только конечные продукты расщепления белков, углеводов, жиров, но и витаминов, гормонов, макро- и микроэлементов. Не исключено, что некоторые энзимы и ряд субстанций стимулируют развитие и созревание паразитов. Использование витаминов гельминтами бывает достаточно велико. Например, О. И. Русович (1990) установила, что в 1 г сырой ткани зрелых члеников мониезий концентрация витамина В12 достигала 4,988±0,21 нг — примерно столько же, сколько в крови здоровых ягнят, — 4,318±0,05 нг/мл. 24
Инокуляторное воздействие паразитов направлено на то, чтобы личинки многих гельминтов (диктиокаулюсы, аскариды, стронгйлиды, стронгилоидесы), насекомых (подкожные и желудочные оводы) или же молодые фасциолы, парамфистомумы и др. в период тканевой миграции переносили во многие органы и ткани хозяина различные виды микроорганизмов. Установлено обсеменение при миграции паренхиматозных органов животных личинками эхинококкусов. Многие двукрылые насекомые при сосании крови инокулируют в организм здоровых животных возбудителей инфекционных болезней. Эймерии в период шизогонального развития, разрушая эпителиальные клетки кишечника, открывают микроорганизмам доступ в глубьлежащие ткани хозяина. В результате организм обсеменяется различными микробами, что нередко осложняет течение инвазионных и инфекционных болезней.