

11.3. Механіка й енергетика скорочення
Амплітудно-часова характеристика скорочення відображає активність скоротливого апарату м'яза. Однак на розвиток сили вкорочення м'яза за його активності сильно впливають також пасивні властивості його елементів. Дійсно, якщо закріпити кінці розслабленого м'яза в міограф за довжини у стані спокою, то, природно, ніякого напруження в ньому не буде. Але якщо потім, від'єднавши один кінець, розтягувати м'яз, то в ньому з'явиться певне пасивне напруження. Зі збільшенням натягу це напруження зростатиме, але не лінійно, як це відбувається при розтягуванні гуми. Відкладаючи на графіку залежність напруження м'яза від його довжини, отримаємо криву напруження спокою (рис. 11.6, а). Отже, м'яз у спокої має деяку пружність. І ця властивість м'яза не залежить ні від його стану (спокій, активність), ні від фармакологічних впливів.
Структурною основою пружності м'яза є: поперечні містки; Z-диски саркомерів; ділянки прикріплення кінців міофібрил до сухожильних елементів м'язового волокна; зовнішні (сухожильні, колагенові та інші сполучнотканинні) елементи м'яза та його волокон; місця прикріплення м'язів до кісток або до приладу в дослідах на ізольованих м'язах; поздовжня система саркоплазматичного ретикулума; сарколема м'язового волокна. Перші п'ять елементів створюють так звану послідовну пружність м'яза, оскільки вони з'єднані послідовно зі скоротливим елементом, інші ж з'єднані паралельно й утворюють паралельну пружність м'яза. Таким чином, механічна модель м'яза складається з трьох елементів: активного скоротливого й двох пасивних, послідовного й паралельного, пружних елементів (рис. 11.6, б).
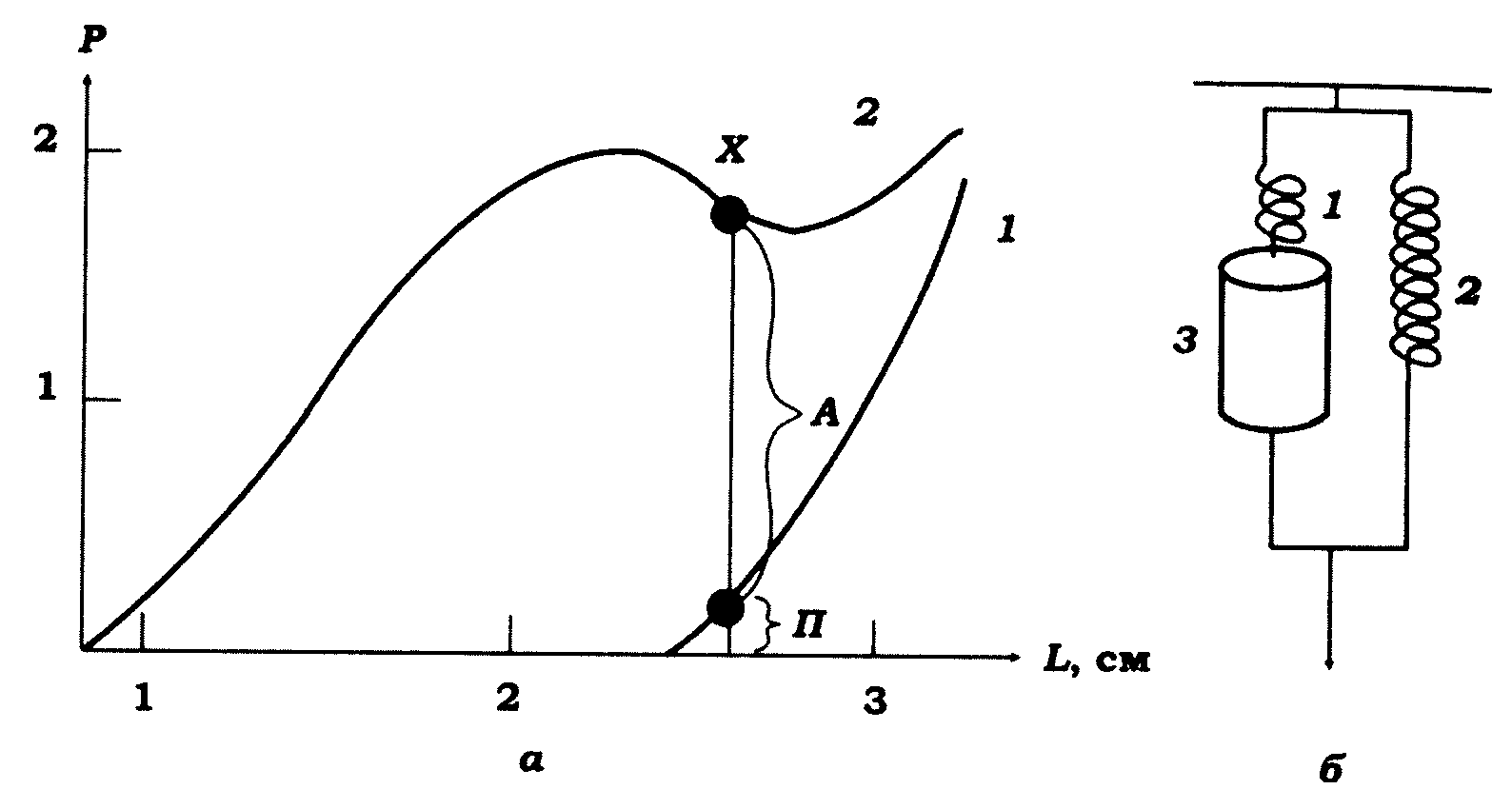
Рис. 11.6. Активні й пасиви! механічні характеристики
поперечносмугастого м'яза: а - залежність між напруженням Р
і довжиною L м'яза (крива довжина - сила); 1 - крива пасивного
напруження; 2 - крива активного ізометричного напруження.
У точці X загальне ізометричне напруження складається з активного (А)
і пасивного (П) компонентів (сила Р виражена в Ньютонах);
б - аналогова модель м'яза: 1, 2 - послідовний і паралельний пружні
пасивні елементи; 3 - скоротливий елемент
Скоротливий елемент, що складається з протофібрил і поперечних містків, у розслабленому стані не чинить опору розслабленню м'яза. У цьому стані активові та міозинові протофібрили не зв'язані між собою поперечними містками й можуть вільно переміщатися. Але за активного стану (скорочення) м'яза певна податливість поперечних містків робить свій внесок у величину послідовного пружного елемента м'яза. Оцінити властивості останнього в цілому м'язі досить важко, бо внесок, наприклад, сухожилля в загальну довжину різних м'язів не є однаковим. Так, у м'язах щурів послідовний пружний елемент становить 7 % оптимальної довжини м'яза, а у кравецькому м'язі жаби - лише близько 2 %. Ці відмінності пов'язані з неоднаковим вмістом і різницею у пружних властивостях сухожилля в цих м'язах.
Залежно від того, чи фіксовані нерухомо обидва кінці м'яза чи тільки один, розрізняють відповідно ізометричний та ізотонічний режими скорочення. При ізометричному скороченні розвиток напруження активованого м'яза не супроводжується його вкороченням. Отже, довжина м'яза при ізометричному скороченні залишається сталою. При ізотонічному ж скороченні м'яз укорочується. До вільного кінця м'яза підвішується певний вантаж Р, і тоді добуток його ваги на ступінь укорочення м'яза (висоту підняття вантажу) і є механічною роботою, виконаною м'язом при скороченні.
У природних умовах, як правило, ні один із цих режимів скорочення м'яза в чистому вигляді не зустрічається. М'яз, скорочуючись, одночасно і вкорочується, і розвиває певне напруження. Такий режим скорочення називається проміжним, або ауксотонічним.
Ізометричне скорочення. В ізометричному режимі поодиноке подразнення м'яза електричним струмом (прямокутний імпульс тривалістю 0,2-0,5 мс) викликає швидке наростання напруження м'яза, яке, досягнувши свого максимуму, потім так само швидко спадає до нуля (рис. 11.7).
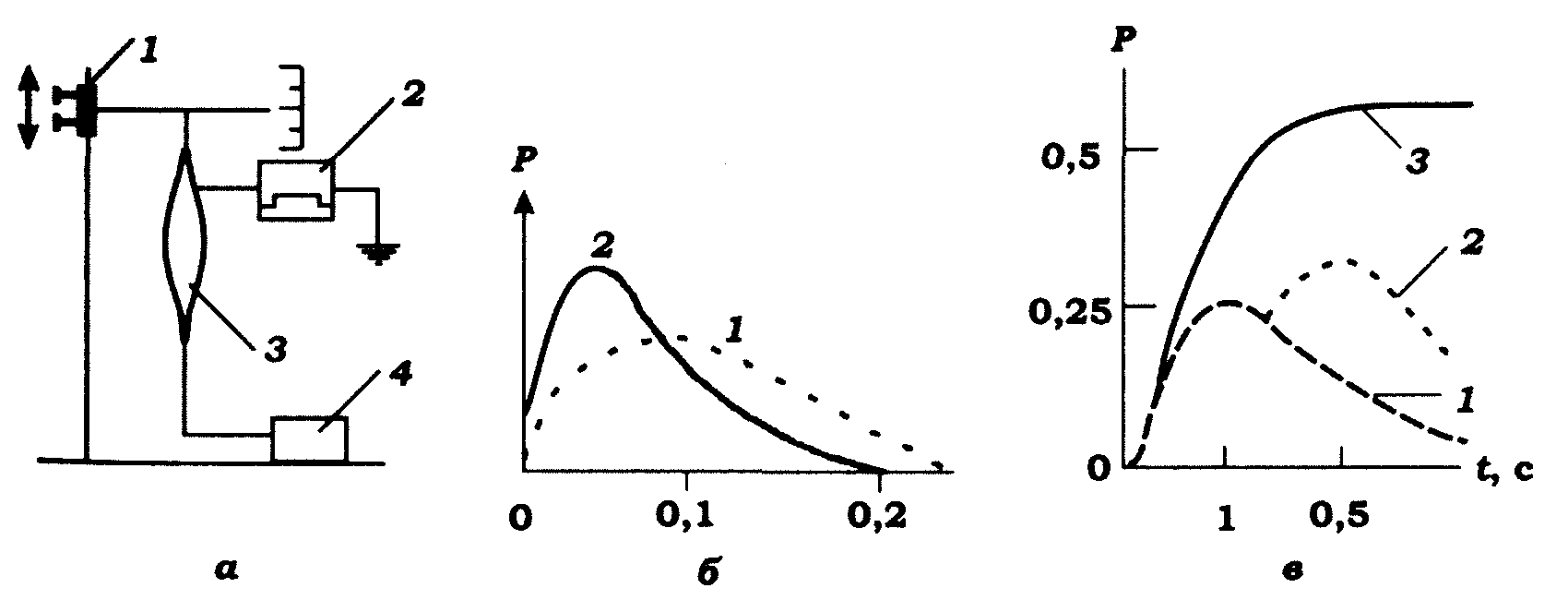
Рис. 11.7. Пристрій для реєстрації ізометричного скорочення й форма
кривих за різних способів стимуляції: а. - елементи пристрою: 1 - пристрій для створення початкового натягу м'яза; 2 - стимулятор; 3 - м'яз; 4 - вимірювач ізометричного напруження м'яза; б - поодиноке
ізометричне скорочення (1) і активний компонент (2) скоротливого
елемента за поодинокого ізометричного скорочення м'язового волокна;
в- підсумовування й злиття поодиноких скорочень (1) у зубчастий (2)
і злитнй (3) тетанус (сила скорочення Р - в Нютонах)
Проте навіть в ізометричних умовах, коли довжина активованого м'яза не змінюється, довжина саркомерів не залишається сталою. За збільшення активного напруження м'яза міозинові голівки скоротливого елемента через поперечні містки, Z-диски, місця кріплення міофібрил тощо надають силу сухожиллям. У результаті послідовний пружний елемент розтягується, а саркомери, навпаки, вкорочуються приблизно на 1 %. Тому навіть в експерименті на ізольованому м'язі скорочення носить до деякої міри ауксотонічний характер, оскільки за рахунок саркомерів відбувається невелике вкорочення м'язових волокон.
Таким чином, розвиток у часі ізометричного напруження м'яза відображає не лише стан активності скоротливого елемента, але й зміну напруження пасивного послідовного пружного елемента (рис. 11.7, б). У цьому зв'язку реально оцінити ступінь активності скоротливого елемента можна тільки шляхом віднімання напруження пружного елемента від загального ізометричного напруження. Із цією метою користуються так званим методом "швидкого звільнення".
Суть методу полягає ось у чому. У різні моменти часу розвитку ізометричного напруження один кінець м'яза швидко звільняється від нерухомої фіксації, завдяки чому м'яз набуває можливості вкорочуватись в ізотонічному режимі. При цьому м'яз має перебувати під меншим навантаженням, ніж ізометричне напруження в момент звільнення. Перехід від одного режиму скорочення до другого супроводжується вкороченням м'яза у дві фази: спочатку швидко, потім повільно. Швидке вкорочення закінчується через кілька мілісекунд, і його пов'язують з укороченням розтягнутого послідовного пружного елемента м'яза.
Наступна повільна фаза відображає вкорочення скоротливого елемента м'яза. Його швидкість залежить лише від інтенсивності активного стану скоротливого елемента в момент швидкого звільнення м'яза, й тому вона є мірою активного стану скоротливого елемента. Після віднімання напруження пружного елемента від загального ізометричного напруження отримаємо криву розвитку активного стану у скоротливому елементі (рис. 11.7, б). Видно, що активний стан після подразнення дуже швидко стає більшим за амплітудою й меншим за тривалістю, ніж повне ізометричне напруження.
Тетанус. Якщо подіяти повторним поодиноким подразненням на м'яз ще до того, як він встигне розслабитись після попереднього ізометричного скорочення, то виникає нове напруження, яке накладається на попереднє, завдяки чому загальне напруження м'яза стає більшим, ніж у разі поодинокого викликаного скорочення (механічна сумація). Багаторазові повторні подразнення з даною частотою також супроводжуються скороченнями, які накладаються одне на одне. У результаті загальне напруження м'яза складається з певної величини тонічного компонента (унаслідок часткової сумації поодиноких скорочень) і фазних скорочень, що повторюються (рис. 11.7, в). Такий тип скорочення називається зубчастим тетанусом.
Зі збільшенням частоти подразнення зубчастий тетанус поступово переходить у суцільний тетанус, коли окремі фазні скорочення вже не виявляються. Амплітуда тетанічного скорочення, особливо за суцільного тетанусу, завжди є значно більшою, ніж амплітуда поодинокого скорочення (рис. 11.7, в). Відношення напруження поодинокого ізометричного скорочення до тетанічного для більшості м'язів кінцівок ссавців становить 0,3-0,4. Для повільних м'язів це відношення є дещо більшим, ніж для швидких.
Поки відсутнє задовільне пояснення значного збільшення амплітуди тетанічного скорочення порівняно з поодинокими скороченнями. За тієї частоти подразнення м'язів, яка необхідна для одержання суцільного тетанусу (25-50 імп/с), амплітуда потенціалів дії м'язових волокон не змінюється, а концентрація вільного кальцію у волокні залишається приблизно такою самою, як і за поодинокого подразнення. Отже, ні потенціали дії, ні внутрішньоклітинна концентрація вільного кальцію не можуть бути причиною збільшення амплітуди тетанічного скорочення.
Причину цього слід шукати, мабуть, у самому скоротливому елементі. Вважають, що при поодинокому подразненні лише частина поперечних містків міозинових протофібрил активується й вступає у зв'язок з актиновими протофібрилами. При повторних подразненнях кількість з'єднаних містків збільшується, і тим самим збільшується сила втягування актинових протофібрил у проміжки між міозиновими протофібрилами.
Зв'язок довжина - напруження при ізометричному скороченні. Цей зв'язок є однією з найважливіших характеристик скоротливого елемента м'яза, який використовується для обґрунтування теорії ковзання. Було виявлено, що для всіх м'язів існує певна оптимальна початкова довжина (L0), за якої ізометричне напруження досягає максимуму (рис. 11.6, а). Ця довжина лежить у межах 100-120 % довжини у спокої, що зазвичай відповідає нормальній довжині м'яза in situ. Зменшення або збільшення довжини м'яза порівняно з L0 завжди супроводжується зниженням тетанічного напруження.
Відкладаючи на осі ординат величину тетанічного напруження (силу), а на осі абсцис – довжину м'яза, отримаємо криву довжина-напруження (рис. 11.6, а). Проте ця крива відображає і активне, і пасивне напруження м'яза. Для того, щоб одержати криву довжина - напруження для скоротливого елемента активного м'яза, необхідно відняти пасивне напруження від загального ізометричного напруження. Не завжди вдається отримати криву для всіх довжин м'яза, бо збільшення його довжини більше 1,3L0 часто викликає незворотні зміни в м'язі, який за цих умов втрачає здатність відповідати скороченнями на подразнення.
Найбільш важливі дані щодо зв'язку довжина-напруження було отримано в дослідах, в яких вивчалася залежність величини тетанічного напруження від довжини саркомера м'язового волокна (рис. 11.8). Проте в ізометричному режимі при тетанічному скороченні саркомери м'язового волокна не однаковою мірою вкорочуються по всій його довжині.
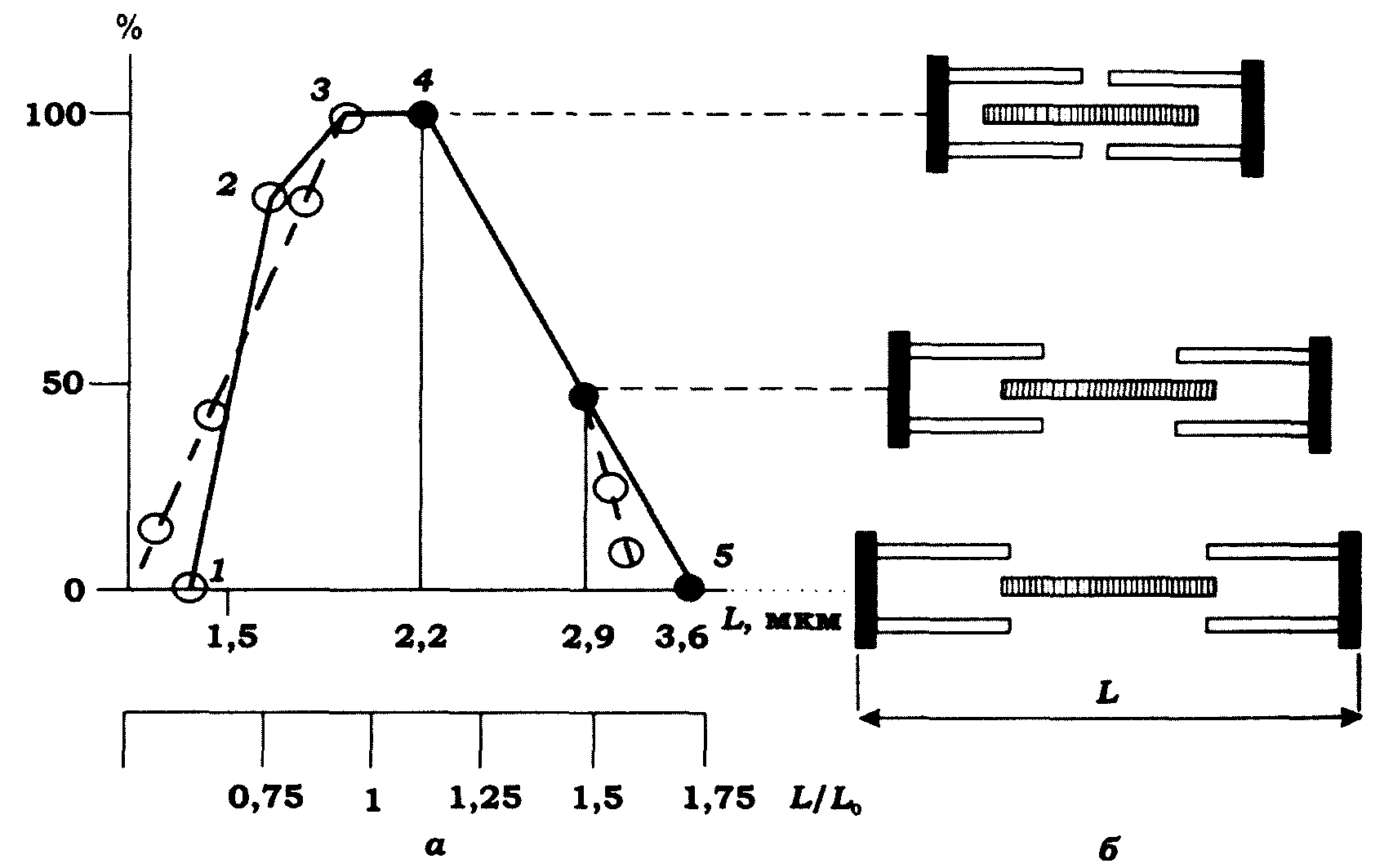
Рис. 11.8. Залежність величини напруження м'язового волокна:
а - від початкової довжини саркомера, % (вісь ординат); б - ступені
перекриття протофібрил. Штрихова лінія - залежність напруження,
що розвивається у кравецькому м'язі жаби, від початкової його
довжини L; L0 - оптимальна довжина м'яза, прийнята за одиницю;
суцільна лінія - залежність довжина - напруження для саркомера
Для уникнення цього необхідно проводити досліди в режимі ізотонічного тетанічного скорочення. На рис. 11.8 показано графічну залежність між довжиною саркомера й напруженням. Видно, що за мінімальної довжини саркомера (до 1,25 мкм, точка 1 на рис. 11.8, а) у відповідь на подразнення напруження не генерується, оскільки взаємне переміщення протофібрил неможливе через початкове повне їх перекриття. Цьому переміщенню заважає також "упирання" протофібрил у Z-диски, оскільки їхня довжина є більшою, ніж початкова довжина саркомера. У міру збільшення довжини саркомера від 1,25 до 1,65 мкм (точка 2) напруження зростає лінійно. Потім крутизна кривої довжина-напруження зменшується, і за довжини саркомера 2-2,2 мкм спостерігається максимальне напруження – стаціонарний режим роботи м'яза (між точками 3 і 4). У такому режимі відбувається максимальне перекриття міозинових й актинових протофібрил і напруження Р не залежить від довжини м'яза.
За подальшого збільшення довжини саркомера відбувається лінійне зменшення напруження, доки за довжини саркомера 3,65 мкм (точка 5 на рис. 11.8, а) воно не стане рівним нулю. Не важко помітити, що за даних довжин саркомера напруження є пропорційним ступеню перекриття товстих і тонких протофібрил (рис. 11.8, б). За довжини саркомера 3,65 мкм і більше актинові протофібрили виходять з проміжків міозинових протофібрил, і тому утворення зв'язку між; ними, а відповідно й взаємне ковзання під дією тягнучої сили поперечних містків стає неможливим.
Порівняємо криву довжина-напруження для саркомера з аналогічною кривою для м'яза (рис. 11.8). За формою ці криві є досить схожими, й максимальне перекриття протофібрил збігається з максимальним ізометричним напруженням (Р0), яке відповідає оптимальній довжині м'яза (L0). Проте крива довжина-напруження для саркомера не є плавною, а має виражені вигини, які відповідають критичним значенням довжини саркомера.
Наведена на рис. 11.8 графічна залежність величини напруження м'язового волокна від ступеня перекриття міозинових і актинових протофібрил втрачається, якщо попередньо надати можливість волокну скоротитись на –3 % і далі проводити вимірювання. У даному разі зменшення напруження при переході від точки 4 до точки 5 іде значно повільніше, ніж показано на рис. 11.8, а. Цей факт неможливо пояснити в межах моделі ковзання протофібрил.
Таким чином, результати досліджень зв'язку довжина-напруження м'яза свідчать про те, що максимальне тетанічне напруження відповідає максимальному перекриттю товстих і тонких протофібрил. Сила м'язів може генеруватись лише при взаємодії актинових і міозинових протофібрил через поперечні містки, утворені голівками молекул міозину.
Ізотонічне скорочення. Під час ізотонічного скорочення відбувається вкорочення м'яза за постійної напруги або навантаження. Для реєстрації цього вкорочення один кінець ізольованого м'яза закріплюється нерухомо, а другий – до вантажу, кінець якого перемішується на величину, пропорційну ступеню вкорочення м'яза (рис. 11.9).
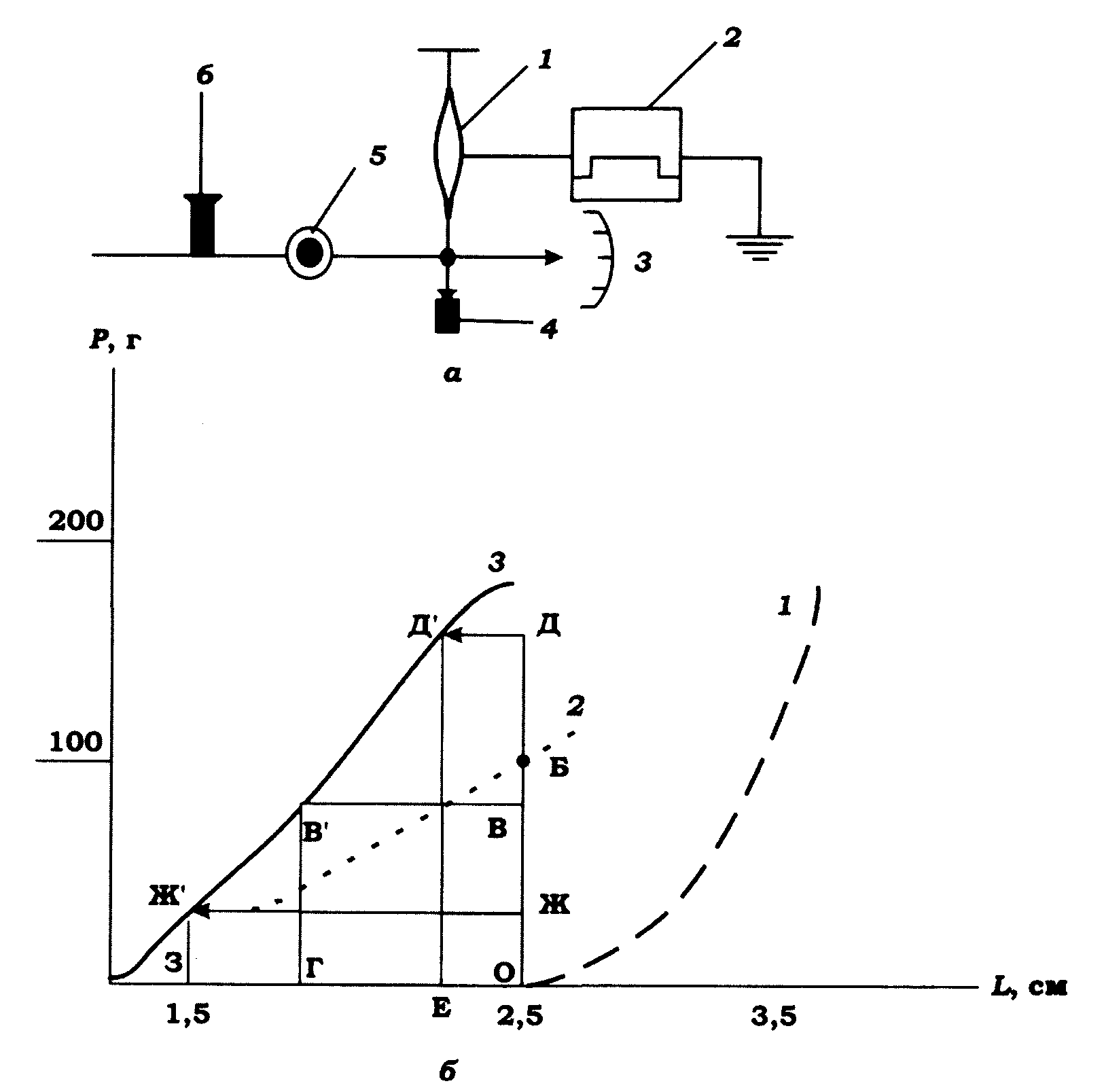
Рис. 11.9. Ізотонічне скорочення м'яза: а - пристрій для реєстрації
ізотонічного скорочення: 1 - м'яз; 2 - стимулятор; 3 - покажчик довжини м'яза;
4 - навантаження; 5 - вісь кріплення навантаження;
6 - гвинт, обмежувальний натяг м'яза; б - залежність
між навантаженням Р і довжиною L м'яза: 1 - крива пасивного
напруження (початкова довжина м'яза 2,5 см); 2 - крива ізотонічних
максимумів; 3 - крива постнавантажувальних ізотонічних максимумів
Подібні дослідження показали, що за однакової початкової довжини м'яза: а) ступінь і швидкість укорочення є тим більшими, чим менше навантаження; б) укорочення досягає свого максимуму тим швидше, чим більше навантаження; в) чим більшим є навантаження, тим пізніше після подразнення починається вкорочення і тим раніше воно закінчується. Ці особливості ізотонічного скорочення м'яза пояснюються таким чином. У відповідь на поодиноке подразнення м'яз спочатку скорочується в ізометричному режимі. Водночас скоротливий елемент, вкорочуючись, розтягує пасивний послідовний пружний елемент. Після того як ізометрична сила м'яза (активне напруження), зростаючи, досягає величини навантаження, м'яз починає вкорочуватись, але вже в ізотонічному режимі, піднімаючи вантаж. Чим більшим буде навантаження, тим меншим – компонент ізотонічного вкорочення м'яза. І коли навантаження досягне значення максимального ізометричного напруження, ніякого вкорочення м'яза не відбуватиметься. Таким чином, підібравши величину навантаження, можна за даної довжини м'яза визначити його максимальне ізометричне напруження (Р0).
Закінчення вкорочення м'яза ще не означає повного завершення розслаблення, бо залишкова релаксація його продовжується в ізометричному режимі, і вона є тим більшою, чим більшим є навантаження. При ізотонічному скороченні м'яз, вкорочуючись, піднімає вантаж і, таким чином, виконує механічну роботу. Величина цієї роботи дорівнює добутку ступеня вкорочення м'яза (ΔL,) на масу вантажу, що піднімається (Р): W = РΔL .
Криву вкорочення-навантаження можна одержати двома шляхами. У першому випадку м'яз попередньо розтягується під дією навантаження, і отже, початкова його довжина варіює залежно від маси вантажу. Це показано на рис. 11.9 кривою напруження спокою 1. Припустимо, що при навантаженні 120 г на кривій 1 довжина м'яза відповідає точці А. При тетанічному подразненні м'яз вкорочується до довжини, показаної на рис. 11.9 точкою Б. Підвішуючи до м'яза різні вантажі, отримаємо криву 2 ізотонічних максимумів.
У другому випадку досліджується вкорочення м'яза за різних навантажень, але початкова довжина м'яза зберігається сталою. Це метод постнавантажувального скорочення. Пасивному розтяганню м'яза під впливом навантаження запобігає стопорний гвинт, як це показано на рис. 11.9, а. Таким чином отримують криву 3 постнавантажувальних ізотонічних максимумів. При кожному навантаженні в м'язі у відповідь на подразнення розвивається спочатку ізометричне напруження. Коли останнє досягає маси вантажу, м'яз починає вкорочуватись, виконуючи роботу. Як видно з рис. 11.9, при навантаженні, наприклад, 90 г виконана м'язом робота дорівнюватиме площі фігури ОВВТ. При навантаженнях 160 і 30 г робота м'яза дорівнюватиме площі фігур ОДД'Е і ОЖЖ'З, відповідно. Площа ОВВТ є більшою, ніж площа ОДД'Е або ОЖЖ'З.
Отже, максимальну роботу м'яз виконує за помірного й середнього навантаження. Збільшення або зменшення навантаження веде до зменшення роботи, а за максимального (рівного ізометричному напруженню) й нульового навантаження робота, виконана м'язом, прямуватиме до нуля.
Важливою характеристикою ізотонічного скорочення є швидкість вкорочення м'яза v. Виявилось, що за однакової початкової довжини м'яза швидкість вкорочення при ізотонічному скороченні є тим більшою, чим меншим є навантаження (рис. 11.10). За нульового навантаження швидкість укорочення м'яза є максимальною, а за максимального навантаження (коли воно дорівнює максимальному ізометричному напруженню – Р0) вкорочення м'яза відсутнє, а тому й швидкість вкорочення дорівнює нулю.
Якщо відкладати по осі абсцис навантаження, а по осі ординат швидкість вкорочення м'яза в ізотонічному режимі скорочення, одержимо криву сила (навантаження)- швидкість, яка має форму гіперболи (рис. 11.10).
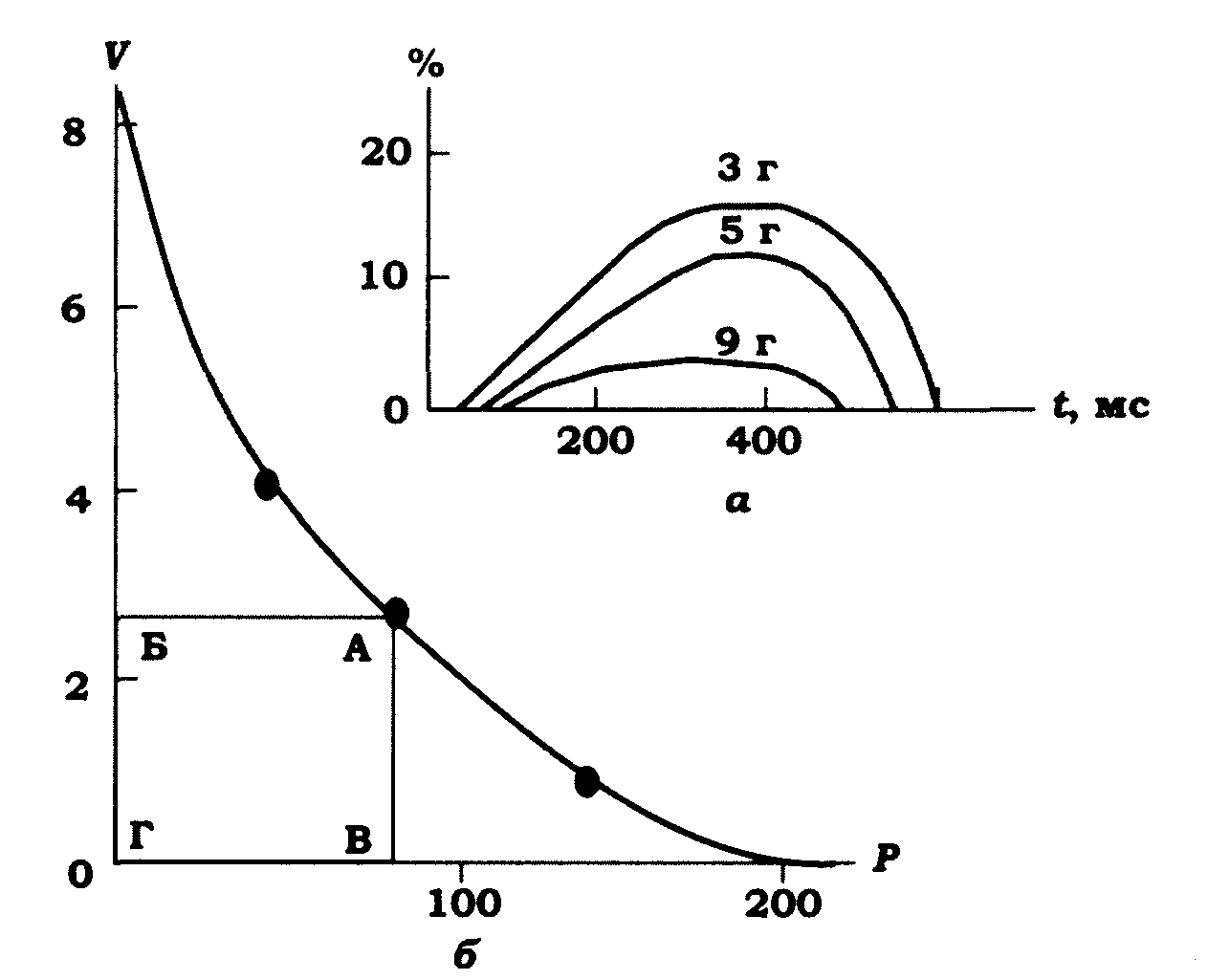
Рис. 11.10. Поодиноке ізотонічне вкорочення й крива сила - швидкість;
а - поодиноке ізотонічне вкорочення м'яза за різних навантажень Р
(ізотонічне вкорочення, % від початкової довжини); б- залежність
швидкості v вкорочення м'яза від величини навантаження Р
за ізотонічного скорочення (крива сила - швидкість), м • с-1.
Площа АБВГ - оптимальна потужність м'яза за швидкості вкорочення 2,5 м • с-1
Оскільки вкорочення послідовно з'єднаних між собою саркомерів м'язового волокна підсумовуються, то при скороченні довгі м'язові волокна вкорочуватимуться швидше, ніж короткі. Наприклад, кравецький м'яз жаби вкорочується зі швидкістю 0,2 м/с, і кожен саркомер довжиною 2 мкм вкорочується на 1 мкм за 50 мс. Більш довгі м'язи кінцівок великих тварин вкорочуються зі швидкістю 8 м/с і більше.
Енергетика скорочення. Під час скорочення м'яза хімічна енергія гідролізу АТФ перетворюється в механічну й теплову.
За допомогою чутливих термопар А. Хілл уперше виміряв малі кількості виділеного тепла. При поодинокому ізометричному скороченні м'яза виділяється енергія, що дорівнює 12,54 Дж • кг-1. В ізотонічному режимі скорочення загальну зміну енергії в м'язі можна розбити на кілька компонентів. Швидке виділення тепла спостерігається на початковій стадії активації скорочення, коли ще відсутнє помітне вкорочення м'яза. Це теплота активації А. Вона виділяється в результаті вивільнення іонів Са2+ із саркоплазматичного ретикулума.
У міру вкорочення м'яза виділяється основна кількість теплоти, яка зветься теплотою вкорочення Qв. Невелика частина теплоти виділяється або поглинається при розслабленні м'яза ±Qр. З урахуванням виконуваної механічної роботи W загальний енергетичний баланс Е для м'яза становитиме:
![]()
Хілл встановив важливий факт, що при ізотонічному скороченні теплота вкорочення Qв є пропорційною величині вкорочення м'яза ΔL:
![]()
де а – коефіцієнт теплоти вкорочення. Оскільки W = РΔL, (11.1) можна записати в такому вигляді:
![]()
Продиференціювавши в часі (11.3), знаходимо потужність N м'яза:
![]()
де V – швидкість вкорочення м яза.
За однакової зміни довжини м'яза швидкість виходу на новий ізометричний рівень зменшується зі збільшенням навантаження Р, при цьому надлишкова потужність порівняно з ізометричною пропорційна Р0 – Р. На основі цього Хілл одержав рівняння:
![]()
де b - константа, Р0 - максимальне ізометричне напруження м'яза. Порівнюючи між собою вирази (11.4) і (11.5), отримаємо відоме рівняння Хілла:
![]()
Дане рівняння встановлює зв'язок між напруженням і швидкістю вкорочення м'яза в ізотонічних умовах, і воно є справедливим у стаціонарному режимі роботи м'яза.
Перетворимо (11.6) таким чином:
![]()
і потім перенесемо змінні величини Р і V в ліву частину рівняння:
![]()
Додамо до лівої та правої частини рівняння добуток сталих Хілла ab:
![]()
У підсумку реформоване рівняння Хілла матиме такий вигляд:
![]()
Залежність, що випливає з (11.7), підтверджено експериментальними біомеханічними дослідженнями (рис. 11.10). При навантаженні Р = Ро м'яз взагалі не скорочується і v = 0. Водночас ненавантажений м'яз (Р = 0) скорочується з максимальною швидкістю v0 і
![]()
Для кравецького м'яза жаби a/P0 = 0,25. Таке саме значення було одержано для поперечносмугастих м'язів щура, кішки, інших тварин.
У більш точних експериментах Хілл встановив, що параметр а не є константою, а залежить від Р.
![]()