

21. Лекарственные средства, стимулирующие эритропоэз
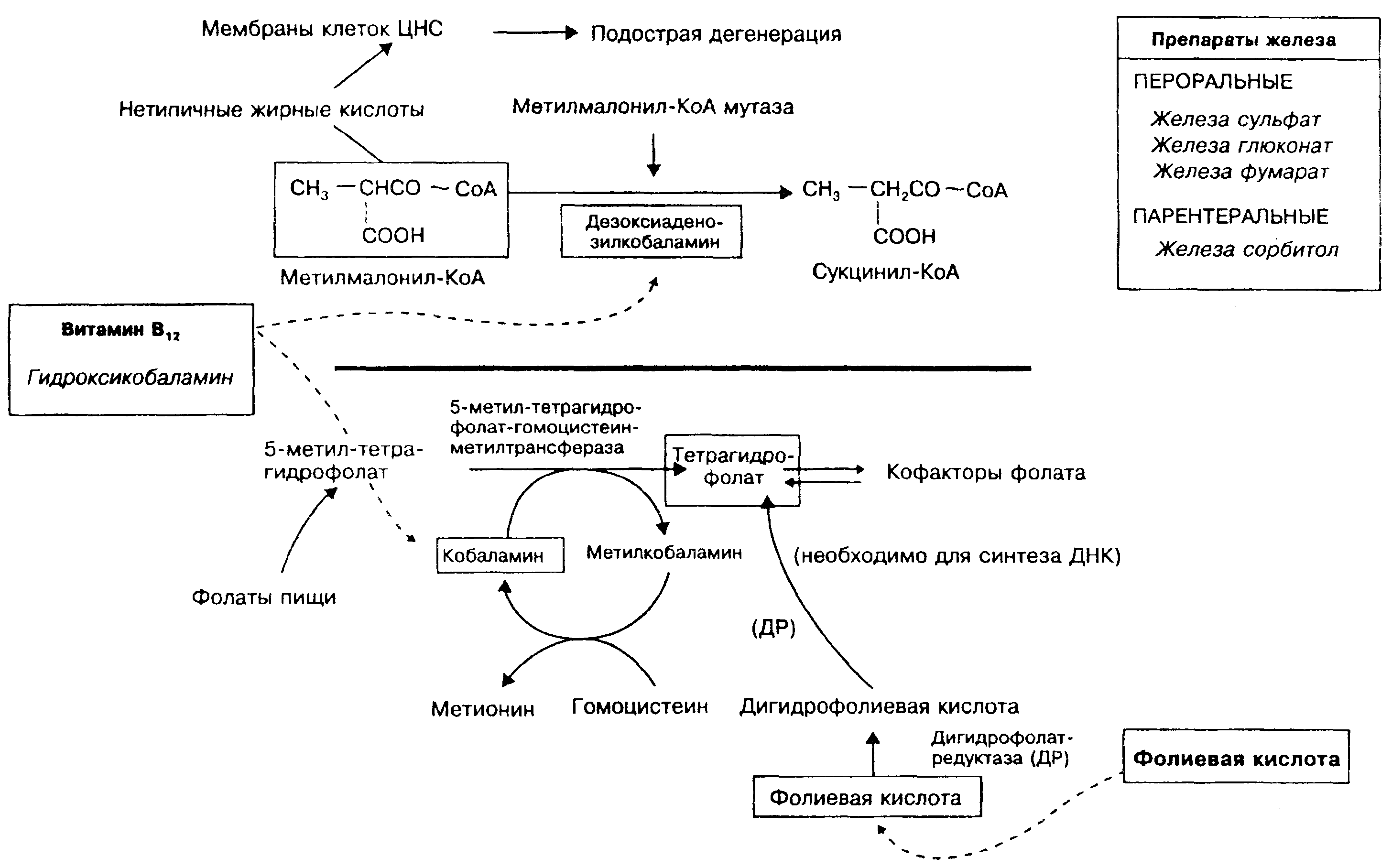
Для нормального эритропоэза необходимы ионы железа, витамин В,; и фолиевая кислота, дефицит которых приводит к развитию анемии. Эритропоэз регулируется эритропоэтином — гормоном, образующимся преимущественно в почках (в антс-натальном периоде синтезируется также в печени). При хронических заболеваниях почек из-за сниженной продукции эрит-ропоэтина часто наблюдается развитие анемии.
Железо необходимо для синтеза гемоглобина. Дефицит железа в организме приводит к образованию эритроцитов с малым содержанием гемоглобина (т.е. к развитию микроцитар-ной гипохромной анемии). При гипохромной анемии, связанной с хронической кровопотерсй, беременностью (плод забирает железо у матери), различными заболеваниями кишечника (уменьшается всасывание железа из кишечника), у недоношенных детей (малые запасы железа в организме при рождении) применяют препараты железа (показаны в верхней части рисунка справа).
Основными побочными эффектами перорального применения препаратов железа являются частые желудочно-кишечные расстройства. Лечение железодсфицитной анемии продолжают до нормализации уровня гемоглобина в крови, затем в течение нескольких месяцев применяют препараты железа в меньших дозах для восстановления его запасов в депо. Дети высоко чувствительны к токсическому действию железа, ребёнок может погибнуть даже при введении такой небольшой дозы, как 1 г железа сульфата. При псрсдозировкс препаратов железа пе-рорально или парентерально вводят дефероксамин (дссферал), который образует с железом хслатные комплексы.
Витамин В |2 и фолиевая кислота необходимы для синтеза в организме ДНК. Дефицит любого из этих витаминов в организме может нарушать эритропоэз (приводить к развитию мегалобластной анемии). Дефицит витамина В\^ в организме вызывает также дегенеративные изменения в центральной нервной системе. Развитие В^-дефицитной анемии связано с нарушением синтеза тетрагидрофолиевой кислоты (показано в нижней части рисунка), а дегенеративные изменения в центральной нервной системе при дефиците витамина В|2 возникают вследствие накопления в клетках головного мозга мстилмало-нил-коэнзима А (показано в верхней части рисунка).
Дефицит витамина b|^ (В^-дефицитная анемия, псрниии-озная анемия, анемия Аддисона-Бирмсра) в организме наблюдается при нарушении его всасывания в желудочно-кишеч-ном тракте вследствие пониженной (или при отсутствии) секреции внутреннего фактора Касла (например, после резекции желудка), а также при различных заболеваниях тонкого кишечника. При В12-дефицитной анемии нарушен синтез ДНК. угнетаются процессы деления клеток, но синтез РНК и белка продолжается, в результате образуются крупные хрупкие эритроциты. При пернициозной анемии витамин В^ применяют и качестве средства заместительной терапии пожизненно инъек-ционно (на рисунке слева). Пероральное применение препарата эффекта не дает. Препаратом выбора для проведения заместительной терапии является оксикобаламин, поскольку он дольше сохраняется в крови, так как более прочно связывается i;
белками плазмы и медленнее выделяется с мочой, чем циано-кобаламин.
Дефицит фолиевой кислоты, который может возникать при беременности (вследствие повышения потребности вфолатах), а также при синдроме малабсорбции (например, при стеатор-рее или спру), приводит к развитию макроцитарной анемии. При фолиеводсфицитной анемии используют пероральное введение фолиевой кислоты.
Железо
Гем, содержащий в своём ядре железо, в комбинации с соответствующими глобиновыми цепями образует протеин — гемоглобин. В составе гемоглобина находится более 90% недепонированного железа (около 2,3 г). Меньшее количество железа (около 1 г) в форме ферритина и гемосидерина депонируется в макрофагах, в селезёнке, печени и костном мозге.
Абсорбция. Железо всасывается в двенадцатиперстной кишке и в проксимальном отделе тощей кишки. В норме всасывается только 5-10% принятого с пищей железа (около 0,5-1 мг в день), но при снижении запасов железа в организме абсорбция его возрастает. Абсорбции подвергаются лишь ионы Fe2^ за счёт активного транспорта. В плазме крови железо находится в комплексе с трансферрином, являющимся р-глобулином. Специального механизма экскреции железа из организма нет — его баланс регулируется только процессами абсорбции.
Препараты железа
Перорально применяют только препараты двухвалентного железа (Fe ), поскольку соединения трёхвалентного железа (Fe^) из кишечника практически не всасываются. При железодефи-цитных состоя ниях для синтеза гемоглобина требуется 50—100 мг железа ежедневно. Поскольку при пероральном применении препаратов железа абсорбироваться может только около 25% принятой дозы, то для быстрой коррекции недостатка железа в организме его необходимо применять в дозе 200—400 мг вдень. При появлении желудоч но-кишечных расстройств (тошнота, боли в животе, понос или запор) возможно снижение дозы железа, однако в этом случае дефицит железа в организме устраняется медленнее.
При невозможности перорального применения препаратов железа их назначают парентерально, однако это обычно не ускоряет повышение уровня гемоглобина в крови.
Фербитол (железа сорбитол) — это комплекс сорбитола, железа и лимонной кислоты. Препарат вводят глубоко внутримышечно, чтобы не вызвать окрашивания кожи (внутривенно фербитол не используют). Парентеральное применение препаратов железа может привести к развитию анафилактической реакции.
Токсичность железа. Острое отравление железом может наблюдаться у детей при приёме большого количества таблеток, содержащих железо. Отравление препаратами железа характеризуется некротическим гастроэнтеритом с болью, рвотой, кровянистым поносом, позднее возможно развитие коматозного состояния. Даже после видимого улучшения состояния при отравлении железом может развиться ацидоз, кома и возможен летальный исход.
Витамин В,^
Витамин В ]2 (внешний фактор Касла) всасывается из кишечника только в комплексе с внутренним фактором Касла (гли-копротеином, секретируемым париетальными клетками слизистой оболочки желудка). Всасывание витамина b)? происходит в дистальном отделе подвздошной кишки при участии высоко специфичной транспортной системы; в крови витамин В^ транспортируется в связи с транскобаламином II (гликопроте-ином плазмы крови). При пернициозной анемии дефицит внутреннего фактора Касла возникает вследствие продукции ауто-антител к самому гликопротеину или к париетальным клеткам желудка (развивается атрофический гастрит). В центре молекулы витамина В^ находится атом кобальта, который ковалентно связывается с различными лигандами, образуя кобаламины. Активными формами витамина В^ являются метлтобаламин и дезоксиаденоэилкобаламин.
Метилмалонил-КоА-мутаза. Этот фермент при участии де-зоксиаденозилкобаламина способствует превращению метил-малонил-коэнзим А в сукцинил-коэнзим А. При отсутствии витамина В)з эта реакция не идет, поэтому накапливается ме-тилмалонил-коэнзим А. В результате происходит синтез аномальных жирных кислот, которые встраиваются в мембраны нервных клеток и вызывают нарушения их функции, что ярко проявляется при дефиците витамина Bi2.
5-метил-тетрагидрофолат-гомоцистеин метилтрансфераза превращает 5-метил-тетрагидрофолат и гомоцистеин соответственно в тетрагидрофолат и метионин. Одновременно в этой реакции происходит превращение кобаламина в метилкобаламин. При дефиците витамина Bi2 эта реакция угнетается, в результате не происходит превращение 5-метилтетрагидрофолиевой кислоты (основного пищевого и эндогенного фолата) в тетрагидро-фолиевую кислоту, необходимую для синтеза ДНК. Эта реакция связывает метаболизм фолиевой кислоты и витамина В ^ и объясняет тот факт, что применение фолиевой кислоты в больших дозах при дефиците витамина Bi2 может уменьшить выраженность анемии, но не влияет на дегенеративные изменения в нервной системе.
ФоАиевая кислота
Запасы фолатов в организме относительно невелики (5-20 мг), тогда как суточная потребность в них высока, поэтому при нарушении поступления в организм фолиевой кислоты анемия развивается достаточно быстро (в течение 1—6 месяцев). Фоли-евая кислота полностью абсорбируется в проксимальном отделе тощей кишки. Фолаты пищи представлены в основном по-лиглутаматами 5-метилтетрагидрофолиевой кислоты. Попиглу-таматы перед всасыванием подвергаются гидролизу с образованием моноглутаматов 5-метилтетрагидрофолиевой кислоты.
Дефицит фолиевой кислоты в организме наиболее часто связан с недостаточным поступлением фолатов с пищей (в отличие от дефицита витамина Bi2, связанного с нарушением его абсорбции). Приём контрацептивов, фенитоина, изониазида и других препаратов может снижать всасывание фолиевой кислоты из кишечника и вызывать её недостаточность в организме.
Фолиевая кислота и витамин Bi2 не оказывают токсического действия. Следует отметить, что при В^-дефицитной анемии нельзя применять только фолиевую кислоту, так как при этом выраженность анемии уменьшается, но усиливаются неврологические нарушения (могут приобрести необратимый характер).
Эритропоэтин
Синтез гемоглобина и выход эритроцитов в кровеносное русло стимулируют гипоксия или кровопотеря. Эти процессы приводят к повышению содержания в крови эритропоэтина (гликопротеина, содержащего 166 аминокислотных остатков). Эритропоэтин связывается с рецепторами на мембранах стволовых клеток-предшественников эритропоэза в красном костном мозге и повышает транскрипцию фермента, участвующего в синтезе гема. Рекомбинантный Эритропоэтин (эпоэтин) используют внутривенно или подкожно при анемии, связанной с нарушением выработки эритропоэтина при хронической почечной недостаточности.
Аминокислоты
7-аминомасляная кислота (ГАМК) присутствует во всех образованиях центральной нервной системы, в основном в тормозных промежуточных нейронах. ГАМК вызывает быстрое торможение центральных нейронов, воздействуя на постсинапти-ческие ГАМКд-рсиепторы, которые блокируются бикукулли-ном (судорожным ядом). Некоторые ГАМК-рецепторы (ГАМКв) не блокируются бикукуллином, но избирательно активируются баклофеном (парахлорфенил-ГАМК). Многие ГАМКв-рецеп-торы расположены на пресинаптических мембранах нервных окончаний, их активация приводит к уменьшению выделения медиаторов (например, глутамата или самой ГАМК). Баклофен уменьшает высвобождение глутамата в спинном мозге и оказывает антиспастическое действие, что используется для уменьшения спастичности при рассеянном склерозе.
Высвобождаясь из пресинаптических нервных окончаний, аминокислотные неиромелиаторы инактивируются в основном путём обратного захвата.
К ГАМК-ергическим средствам относятся бензодиазепины. барбитураты (см. главу 24). противосудорожные препараты ви-габатрин и. возможно, вальпроаты (см. главу 25).
Глицин — это тормозной медиатор промежуточных нейронов спинного мозга. Глициновые рецепторы блокируются стрихнином, а высвобождение глицина и^ нервных окончаний угнетается столбнячным токсином. Оба вещества вызывают судороги.
Глутамат возбуждает практически все центральные нейроны, взаимодействуя с аминокислотными рецепторами различных типов. Эти рецепторы подразделяются на АМРА (активируются а-амино-3-гидрокси-5-мегил-4-изоксазолпропионовой кислотой), NMDA (активируются ^'-метил-О-аспартатом) и каинатныс (активируются каиновой кислотой). Существует также группа метаботропных рецепторов (связаны с G-белками). Антагонисты NMDA-реиспторов (например, 2-аминофосфоно-валерат) обладают противоэпилептической активностью ^ эксперименте на животных и оказывают благоприятное действие при судорожном синдроме, связанном с нсйрональными нарушениями вследствие значительного высвобождения глутамата. Ламотриджин является противоэпилептическим средством, механизм действия которого связан с угнетением высвобождения глутамата из пресинаптических мембран нейронов (см. главу 25).
Моноамины
Аиетилхолин является в основном возбуждающим медиатором LIHC. Он высвобождается из нервных окончаний мотонейронов нервно-мышечных синапсов и в синапсах коллатеральных аксонов с клетками Реншоу в спинном мозге (это примеры н-холинергичсских синапсов). Возбуждающие эффекты ацетилхолина на центральные нейроны в основном опосрсду-ются через м-холинорецепторы. активация которых приводит к угнетению потенциалзависимого К^тока в мембранах не-овных клеток. Это способствует возбуждению нервных клеток и облегчению формирования их ответа на тонические возбуждающие импульсы.
Холинергических нейронов особенно много в базальных ганглиях. Они принимают участие в проведении восходящих кортикальных импульсов и в процессах формирования памяти. М-холиноблокаторы (атропиноподобные средства) могут вызывать нарушения памяти (амнезию). Скополамин (гиосцин) используют в анестезиологии для премедикации в качестве средства, вызывающего амнезию и оказывающего успокаивающее действие (см. главу 23). Центральные эффекты препаратов этой группы используют при морской болезни и паркинсонизме (см. главу 26). При болезни Альцхаимера (форма сенильной демен-ции, лечение которой в настоящее время практически не эффективно) отмечается снижение числа холинергнческих нейронов и ухудшение памяти. Такрин — это антихолинэстеразное средство, которое оказывает благоприятное действие при болезни Альцхаймера (у 15% пациентов), но обладает выраженной гепатотоксичностью.
Катехоламины (при локальном нанесении на центральные нейроны) оказывают преимущественно тормозной эффект.
Дофаминергические пути проецируются от субстанции «ниг-ро» в средний мозг к базальным ганглиям и из среднего мозга в лимбическую кору и другие лимбические структуры. Тубсро-инфундибулярный дофаминергический путь участвует в регуляции высвобождения пролактина, а нигростриатный путь — в управлении произвольными движениями, его поражение приводит к развитию паркинсонизма. Мезолимбический дофаминергический путь чрезмерно активируется при шизофрении. однако причины этого неизвестны. Агонисты аофамина используют при паркинсонизме (см. главу 26), а антагонисты (нейро-лептики) применяют при шизофрении (см. главу 27). Дофами-новые рецепторы находятся в хсморсцепторной триггсрной (пусковой) зоне, поэтому антагонисты дофаминовых рецепторов оказывают противорвотное действие (см. главу 30).
Норадреналинергические нейроны встречаются в нескольких образованиях ствола головного мозга. Больше всего этих нейронов находится в голубом пятне (locus coeruleus) в мосту, от которого аксоны нейронов проецируются на всю дорсальную часть переднего мозга, особенно в кору головного мозга и гип-покамп. Большое количество норадрснергических волокон находится в гиппокампе. Норадрсналин и дофамин в лимбичес-ких структурах переднего мозга (особенно nucleus accumbens) могут участвовать в формировании чувства «удовольствия», что имеет значение в развитии лекарственной зависимости (см. главу 31). Нарушение адренергических функций может быть причиной развития депрессии (см. главу 28).
Серотонин (5-гидрокситриптамин, 5-НТ) находится в основном в телах нейронов ядер шва ствола мозга, которые посылают волокна в передний мозг, а также к вентральным и дорсальным рогам спинного мозга. Последние могут модулировать болевые импульсы (см. главу 29). Нарушения в ссротонинерги-ческой системе, также как и в норадреналинергической системе, могут приводить к возникновению депрессии. 5-НТз-рсцеп-торы локализованы п триггерной зоне рвотного центра, поэтому антагонисты этих рецепторов обладают антиэметическим (противорвотным) действием. 5-НТ]о-рецепторы находятся в сосудах головного мозга, агонист этих рецепторов суматриптан облегчает течение мигрени, суживая сосуды головного мозга. чрезмерно расширенные во время приступа.
Неиропептиды образуют самую многочисленную группу центральных нейромедиаторов, функция которых малоизвестна. Субстанция Р и энксфалины предположительно участвуют в проведении болевых импульсов (см. главу 29).
Оксид азота (NO). Синтез NO осуществляется в 1—2% нейронов многих областей головного мозга — таких, как кора головного мозга, гиппокамп и стриатум. Образование NO происходит совместно с выделением других нейромедиаторов: глутамата, ГАМК, соматостатина, нейропептида Y. NO влияет на процессы высвобождения некоторых нейромедиаторов. Физиологическая роль NO в головном мозге не ясна, однако существуют сведения, что NO может регулировать синаптическую пластичность.