

3.Трансаминирование амк. Аминотрансферазы. Отдельные аминотрансферазы. Реакции. Биохимическое значение трансаминирования амк.
Под трансаминированием подразумевают реакции межмолекулярного переноса аминогруппы (NH2—) от аминокислоты на α-кетокислоту без промежуточного образования аммиака.
Реакции
трансаминирования являются обратимыми
и, как выяснилось позже, универсальными
для всех живых организмов. Эти реакции
протекают при участии аминотрансфераз.
Теоретически
реакции трансаминиро-вания возможны
между любой амино- и кетокислотой, однако
наиболее интенсивно они протекают в
том случае, когда один из партнеров
представлен дикарбоновой амино- или
кетокислотой. Все трансаминазы (как и
декарбоксилазы аминокислот) содержат
один и тот же кофермент – пиридоксальфосфат.
Для реакций трансаминирования харак
-терен общий механизм. Специфичность
трансаминаз обеспечивается белковым
компонентом. Ферменты трансаминирования
катализируют перенос NH2-группы не на
α-кетокислоту, а сначала на кофермент
пиридоксаль-фосфат. Образовавшееся
промежуточное соединение (шиффово
основание) подвергается внутримолекулярным
превращениям (лабилизация α-водо-родного
атома, перераспределение энергии связи),
приводящим к освобождению α-кетокислоты
и пиридоксаминфосфата; последний на
второй стадии реакции реагирует с любой
другой α-кетокислотой, что через те же
стадии образования промежуточных
соединений (идущих в обратном направлении)
приводит к синтезу новой аминокислоты
и освобождению пиридоксальфосфата.
Опуская промежуточные стадии образования
шиф-фовых оснований, обе стадии реакции
трансаминирования можно представить
в виде общей схемы:
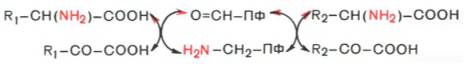
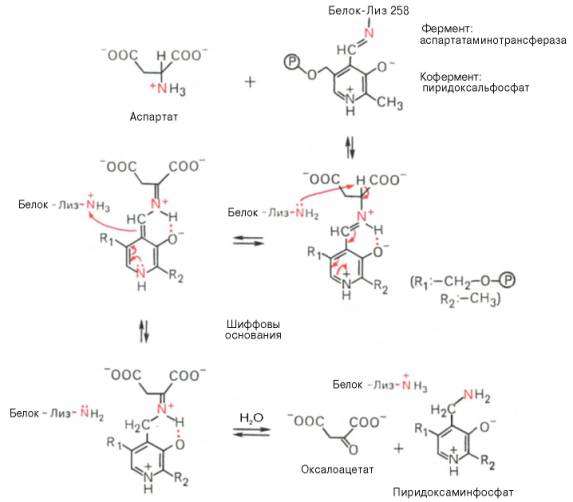
В связи с тем что во всех пиридоксалевых ферментах (включая транс-аминазы) карбонильная группа кофермента (—СНО) оказалась связанной с ε-аминогруппой лизина белковой части, в классический механизм реакции трансаминирования А.Е. Браунштейн и Э. Снелл внесли следующее дополнение. Оказалось, что взаимодействие между субстратом, т.е. L-амино-кислотой (на рисунке – аспартат), и пиридоксальфосфатом происходит не путем конденсации с выделением молекулы воды, а путем реакции замещения, при которой NH2-группа субстрата вытесняет ε-NН2-группу лизина в молекуле ферментного белка, что приводит к формированию пиридоксальфосфатного комплекса.
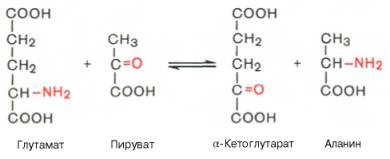
Аминотрансферазы (трансаминазы) — ферменты, катализирующие межмолекулярный перенос аминогруппы от соответствующих аминокислот на a‑кетокислоты (2‑оксокислоты) с образованием новых кето- и аминокислот без образования свободного аммиака, в качестве кофермента используется витамин В6 (пиридоксин). Эти ферменты играют центральную роль в обмене белков, осуществляя окислительное дезаминирование аминокислот опосредованно через глутаминовую кислоту. Образующаяся глутаминовая кислота дезаминируется глутаматдегидрогеназой с освобождением свободного аммиака и 2‑оксоглутаровой кислоты. В организме человека наибольшее значение имеют две аминотрансферазы: аспартатаминотрансфераза (АСТ или АсАТ) (L‑аспартат:2‑оксоглутарат-аминотрансфераза, КФ 2.6.1.1.) и аланинаминотрансфераза (АЛТ или АлАТ), (L‑аланин:2‑оксоглутарат-аминотрансфераза, КФ 2.6.1.2.). В клинической практике чаще всего определяют именно активность этих двух ферментов. Существует также другое название указанных ферментов: для АСТ — глутаматоксалоацетатаминотрансфераза (ГОАТ), для АЛТ — глутаматпируватаминотрансфераза (ГПАТ). Ниже приведены реакции, катализируемые этими ферментами:
2-Оксоглутарат + Аспартат ↔ Глутамат + Оксалоацетат
2-Оксоглутарат + Аланин ↔ Глутамат + Пируват
Наибольшая активность АСТ обнаружена в миокарде, затем в порядке убывания в печени, скелетных мышцах, головном мозге, почках. Активность фермента в миокарде в 10000 раз выше, чем в сыворотке крови. Фермент является димером, имеет изоферменты: положительно заряженный митохондриальный с ММ=93 кД и отрицательно заряженный цитозольный с ММ=92 кД. Активность АЛТ максимальна в печени, среди других органов убывает в последовательности: поджелудочная железа, сердце, скелетные мышцы, селезенка, легкие. Фермент также имеет цитозольный и митохондриальный изоферменты, однако последний содержится в минимальном количестве и нестабилен. Избирательная тканевая локализация позволяет считать трансаминазы маркерными ферментами: АСТ для миокарда, АЛТ для печени. Соотношение активности аминотрансфераз позволяет судить о глубине повреждения клеток: АЛТ преимущественно локализована в цитоплазме, АСТ — в цитоплазме и в митохондриях.
Константа равновесия ферментативной реакции трансаминирования приблизительно равна 1, то есть направление протекания процесса переноса аминогруппы определяется концентрацией субстратов и продуктов в клетке. Одна и та же реакция трансаминирования может протекать в разных направлениях в разных частях клетки. Следовательно трансаминирование служит как для анаболизма, так и для катаболизма аминокислот, то есть является амфиболическим процессом.
Лизин и треонин не участвуют в реакциях трансаминирования.
Продуктами чаще всего являются аланин, аспарагин и глутамат, так как соответствующие им кетокислоты образуются в процессе метаболизма углеводов. Трансаминирование играет важную роль в процессах мочевинообразования, глюконеогенеза, путях образования новых аминокислот. Трансаминирование аминокислот с образованием глутаминовой кислоты в сочетании с ёё дезаминированием НАД(Ф)-зависимой глутаматдегидрогеназой называется непрямым дезаминированием аминокислот (трансдезаминирование).