

Билет 13. Фолиевая кислота. Биологическая роль. Пути поступления в организм. Коферментная функция.
Фолиевая кислота — водорастворимый витамин B9 необходимый для роста и развития кровеносной и иммунной систем.
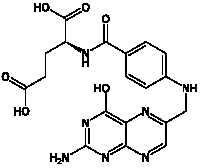
Наряду с фолиевой кислотой к витаминам относятся и её производные, в том числе ди-, три-, полиглутаматы и другие. Все такие производные вместе с фолиевой кислотой объединяются под названием фолацин. Недостаток фолиевой кислоты может вызвать мегалобластную анемию у взрослых, а при беременности повышает риск развития дефектов нервной трубки. Фолиевая кислота необходима для создания и поддержания в здоровом состоянии новых клеток, поэтому её наличие особенно важно в периоды быстрого развития организма — на стадии раннего внутриутробного развития и в раннем детстве. Процесс репликации ДНК требует участия фолиевой кислоты, и нарушение этого процесса увеличивает опасность развития раковых опухолей. В первую очередь от нехватки фолиевой кислоты страдает костный мозг, в котором происходит активное деление клеток. Клетки-предшественники эритроцитов, образующиеся в костном мозге, при дефиците фолиевой кислоты увеличиваются в размере, образуя так называемые мегалобласты и приводя к мегалобластной анемии. Основная функция фолиевой кислоты и её производных — перенос одноуглеродных групп, например метильных и формильных, от одних органических соединений к другим. Главная активная форма фолиевой кислоты — тетрагидрофолиевая кислота, образуемая с помощью фермента дигидрофолат редуктазы. Фолиевая кислота метаболически неактивна, но является предшественником коферментов, включающихся в обменные процессы. Важной химической особенностью фолиевой кислоты является способность ее птеридинового кольца к восстановлению путем присоединения 4 водородных атомов в 5, 6, 7 и 8 положениях с образованием тетрагидрофолиевой кислоты (ТГФК).
Наличие в молекуле ТГФК 4 подвижных атомов водорода обусловливает ее участие в некоторых окислительно-восстановительных реакциях в качестве донора электронов. Однако биологическая роль ТГФК определяется в основном наличием в положениях 5 и 10 молекулы активных в химическом отношении атомов азота, способных присоединять одноуглеродные радикалы. Это свойство лежит в основе коферментных функций ТГФК.
Восстановление фолиевой кислоты подробно изучено вначале в химических, а затем в ферментных системах. Превращение фолиевой кислоты в ее тетрагидроформу происходит через промежуточный продукт, дигидрофолиевую кислоту. Источником электронов в этих реакциях могут быть НАДФ-Н или НАД-Н. Восстановление происходит преимущественно в печени согласно следующим уравнениям:
ФК + НАДФ-Н + Н+ ---------- ДГФК + НАДФ
ДГФК + НАДФ-Н + Н+ ------- ТГФК + НАДФ
Тетрагидрофолат — соединение неустойчивое и в присутствии молекулярного кислорода быстро превращается в дигидрофолат. Он окисляется также и ферментативным путем при участии НАДФ. Тетра-гидрофолиевая кислота является биологически активной формой фолатов. Точно установлено, что ее коферментные функции непосредственно связаны с переносом одноуглеродных групп. Первичными источниками одноуглеродного фрагмента в организме могут служить бета-углеродный атом серина, альфа-углеродный атом глицина, 2-й углеродный атом имидазольного кольца гистидина, 2-й углеродный атом индольного кольца триптофана, углерод метальных групп холина, метионина, диметилглицина, а также образующиеся в организме в процессе обмена формальдегид, муравьиная кислота и метанол. Фолиевая кислота и другие витамины. Наиболее тесная функциональная взаимосвязь существует между фолиевой кислотой и витамином B12. Выше были подробно рассмотрены вопросы взаимодействия фолатов и витамина B12 в таких важных реакциях организма, как синтез пуриновых и пиримидиновых оснований, метионина, в обмене гистидина и т. д. Существует предположение, что витамин B12 участвует также в переносе фолатов в клетку и выведении их из нее. Это предположение подтверждается наблюдениями, свидетельствующими о резком падении содержания фолатов в печени у овец при острой недостаточности витамина В12 и кобальта, а также относительно высокой активностью фолатов в сыворотке и низким содержанием их в эритроцитах у больных с недостаточностью витамина B12.
Состояние обмена фолатов зависит также от обеспеченности организма витамином С. Мегалобластическая анемия, часто наблюдаемая при цинге, по-видимому, является результатом нарушения обмена фолатов, обусловленным недостаточностью витамина С. Высказано предположение, что аскорбиновая кислота предохраняет фолатредуктазу от разрушения. На это указывает, в частности, тот факт, что при введении витамина С увеличивается выделение с мочой восстановленных форм фолатов. Косвенным доказательством взаимодействия этих витаминов служат также данные о том, что отмечаемые у животных с цингой нарушения окисления тирозина, фенилаланина и падение активности оксидазы п-оксифенилпировиноградной кислоты устраняются в равной степени аскорбиновой и фолиевой кислотами.
Весьма интересны факты об участии в обмене фолатов биотина. Было установлено, что недостаточность биотина приводит к значительному уменьшению общего содержания фолатов в печени и к изменению в соотношении восстановленных форм фолатов. Эти данные показывают, что недостаток биотина ухудшает использование фолатов, нарушая превращение их в активные формы.