

5.2. Термодинамика клетки. Возникновение цели.
Базовой ячейкой любого живого организма является клетка. Более того, можно считать, что жизнь – это способ существования клеток – особого природного объекта, характеризуемого непрерывным циклическим протеканием химических реакций, преимущественно полимеризации и деполимеризации белков и нуклеиновых кислот. С этой точки зрения многоклеточные организмы – это сложные по структуре колонии родственных клеток, объединившихся для совместного выживания.
Не вдаваясь в сущность химических реакций, происходящих в клетке, можно определить следующие их особенности:
Все химические реакции – циклические, т.е. протекают по строго установленному порядку, периодически повторяясь; внутри клетки имеется специальная система обратной связи, следящая за соблюдением повторяемости химических реакций и ее длительным сохранением.
Они сопровождаются непрерывным потоком реагентов – в клетку обязательно должны поступать исходные субстраты химических реакций и из неё должны выводиться конечные продукты этих реакций.
Исходя из первых двух особенностей, клетка представляет собой открытую систему, тепловая энергия в которую входит с помощью высокоэнергетических веществ, а выходит с помощью веществ с малыми свободными энергиями и, частично, путём теплопередачи.
Масса субстратов, входящих в клетку, обязательно превосходит массу выводимых из неё продуктов.
Ниже в §4.9 будут изложены современные данные биологической науки о том, как в природе мог возникнуть такой объект. Сейчас же, не углубляясь в суть происходящих в клетке процессов, рассмотрим, каковы необходимые термодинамические условия её существования. Для этого представим клетку в виде шара с полупрозрачной оболочкой и радиусом R (рисунок 5.1,а[16]).
m2
R
Rmin
Q m1
а б
в
г










Рисунок 5.1
Термодинамическая схема деления клетки: а – потоки вещества и энергии в клетке; б – простейший вариант увеличения поверхности клетки – растягивание её в эллипсоид; в – эволюция эллипсоида в “гантель’; г – деление клетки (завершающая стадия её эволюции)
Химические реакции, протекающие в этом шаре-реакторе, относятся к категории автокаталитических (см § 3.6): по мере их протекания увеличивается масса продуктов, катализирующих реакции между ними и поэтому обуславливающих потребность в увеличении веществ, вовлеченных в эти реакции. Отсюда – непрерывное увеличение клетки в объеме и одновременное пропорциональное увеличение вырабатываемой энтропии. Эта энтропия выводится из клетки в окружающую среду частично в виде тепловой энергии Q, частично в виде низкоэнергетических субпродуктов. Инструментом обмена веществом и энергией клетки с окружающей средой служит оболочка клетки. Таким образом, если рассматривать клетку, как открытую систему, то информация в нее (вернее разность поступающей и выводимой информации), необходимая для компенсации возникающей в процессе химических реакций энтропии, поступает через оболочку.
Ясно, что по мере
роста размеров клетки её оболочка
возрастает пропорционально квадрату
радиуса R,
в то время, как масса её реактивов –
пропорционально кубу радиуса.
Следовательно, количество вовлеченной
в химические реакции материи, а значит,
и вырабатываемой энтропии, возрастает
пропорционально R3,
а объем получаемой информации (порядка),
призванной скомпенсировать эту энтропию
– пропорционально R2.
Например, если бы радиус клетки удвоился,
то энтропия внутри нее возрастала бы в
8 раз, а получаемая информация - только
в 4 раза [16]. При определенном радиусе
клетки её поверхности будет недостаточно,
чтобы обеспечивать протекание
упорядоченных (циклических) реакций в
ней. Заключенный в клетке механизм
сохранения устойчивости – механизм
отрицательной обратной связи – вызовет
дополнительные процессы, которые должны
будут обеспечить необходимый приток
информации, компенсирующий возрастающий
объем энтропии [16]. Наиболее простой из
этих процессов – изменение геометрической
формы клетки. На рисунке 5.1 показана
схема этого процесса. Сначала шар
вытягивается в эллипсоид – наиболее
близкое по форме к шару геометрическое
тело, у которого при том же объёме площадь
поверхности больше. Принципиальное
отличие эллипсоида от шара – наличие
двух центров симметрии – сравните
рисунки 5.1, а и б. Вследствие этого
начинается разделение автокаталитического
процесса на два параллельных, что
приводит в конечном счете к делению
клетки на две дочерние – рисунок 5.1 в,
г. После деления клетки ее объем
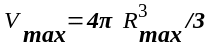 разделился на два минимальных:
разделился на два минимальных:
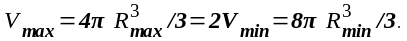
Следовательно,
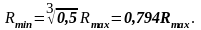 (5.1)
(5.1)
Нетрудно установить,
что энтропия клетки колеблется в пределах
2:1, а получаемая ею извне информации –
в пределах 2:1,3. Таким образом, поддерживается
долговременное протекание циклических
химических реакций. С удвоением
(репликацией) клетки порядок (гармония)
в природе возрастает на
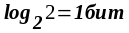 .
Реакции, протекающие в этих дочерних
клетках, в точности повторяют реакции
родительской клетки до последующего
деления на четыре клетки.
.
Реакции, протекающие в этих дочерних
клетках, в точности повторяют реакции
родительской клетки до последующего
деления на четыре клетки.
Как видим, клетки, а вслед за ними все другие биологические объекты вынуждены, чтобы сохранить заложенный в них уровень гармонии, размножаться в геометрической прогрессии. Именно это обстоятельство приводит к качественному усложнению фактора устойчивости – превращению его в фактор цели.
Цель отличается от устойчивости наличием конкурентной борьбы – чтобы сохранить накопленный в них порядок, потомки клетки вынуждены вступать в конфликт друг с другом за обладание исходными продуктами протекающих в них реакций. Если устойчивость связана с процессами внутри диссипативной системы, направленных на восстановление равновесия при стохастических воздействиях окружающей среды, то цель вызывает процессы, направленные на изменение условий в окружающей среде независимо от того, нарушают они внутреннее равновесие системы или нет. Равновесие внутри биологической системы нарушается не столько из-за воздействия внешних факторов, сколько в результате активных внутренних процессов в ней.
В связи с разнообразием условий существования живых организмов (биологических систем) и их самих цели могут отличаться друг от друга. Но, несмотря на такие различия, все они сводятся к главной всеобщей цели – каждая клетка, каждый многоклеточный живой организм стремятся сохранить и приумножить накопленный в них порядок. Эта цель распадается на две подцели – самосохранение, т.е. сохранение самого живого организма, и воспроизведение себе подобного потомства. Обе эти подцели находятся в сложной взаимозависимости друг от друга. У высокоразвитых организмов главной подцелью является самосохранение. У менее развитых организмов на первый план выходит вторая подцель. В частности, трудность борьбы с некоторыми насекомыми, например, мухами или комарами, связано не столько с тем, что нельзя ликвидировать их самих, сколько с невозможностью борьбы с их потомством.
Генезис устойчивости, как фактора существования диссипативных и управляемых неживых систем, в цель – главный фактор сохранения биологической системы – дал мощный импульс к эволюции – возникновению сверхсложных, высокоинформативных объектов.
В конце 18 столетия английский экономист Т.Р.Мальтус [2,12], изучая воздействие этого важнейшего биологического закона на различные популяции, пришел к печальному выводу о неминуемой гибели части их потомства. По сути дела, мы сталкиваемся здесь с той же проблемой, которая привела к мысли о гибели природы в результате неограниченного нарастания энтропии, только на более высоком уровне сложности. Здесь также действует термодинамический закон необратимости во времени естественных процессов. И так же, как в микромире тенденция к хаосу порождает возникновение структур – систем со сложным порядком, так и в биологии тенденция к неограниченному возрастанию определенного биологического вида порождает возникновение более развитых, более сложных организмов.
В отличие от открытых систем в неживой природе, которые могут длительно устойчиво существовать без изменения количества упорядоченности, живые системы не могут длительно существовать без непрерывного накопления гармонии и, следовательно, усложнения структур. В этом смысле удобно сравнение полета самолета и воздушного шара. Если нахождение на высоте последнего в неподвижном состоянии возможно сколь угодно долго, то первый должен непрерывно двигаться, причем с достаточно большой скоростью.
Ещё раз подчеркнём, что, по сути дела, проведённый анализ термодинамики клетки иллюстрирует генезис фактора цели – его происхождение от фактора устойчивости. Действительно, клетка делится потому, что заложенная в ней обратная связь отслеживает разность между возникающей в процессе химических реакций энтропией и поступающей извне информацией – как только она превышает некоторую норму, включается механизм деления клетки. Как видим, никакой качественной разницы с системой регулирования температуры холодильника нет (см. §4.4). Однако возникающее в результате явление – деление клетки – создаёт в природе новую ситуацию: накопление порядка и, как результат, качественный скачок от устойчивости – сохранения порядка – к цели – его умножению.
Цель – это внутренняя причина возникновения процесса. Например, движение воды в реке обусловлено ее накоплением в районе истока – внешняя причина, а движение мухи – ее желанием приобрести пищу, т.е. внутренняя. Строго говоря, цель – это совокупность причин, возникших миллионы лет назад и записанных в наследственной памяти животных.
Таким образом, появление цели, как фактора накопления гармонии внутри систем с упорядоченной структурой, означает ее новый, более высокий уровень. Исходной причиной, породившей цель, является устойчивость систем обратной связи [2]. Разница заключается в том, что устойчивость обеспечивает сохранение передачи порядка от органа памяти исполнительному органу, а цель – сохранение порядка в самом органе памяти, даже при условии, что сам исполнительный орган может быть разрушен. Носителем цели является орган управления живым организмом. Этот орган не только сохраняет ранее приобретенную гармонию, не только обеспечивает устойчивую передачу этой гармонии исполнительному элементу организма, но и стремится накопить более высокую степень упорядоченности.
В конечном счете, именно наличие цели во всех живых организмах и явилось тем "конструктором", который "разработал" и "осуществил" фантастические "изделия" живой природы.
Все, что сказано в этом параграфе, имеет феноменологический характер, т.е. объясняет общее течение процесса в клетке, не раскрывая его механизма. Последнее будет сделано в следующих параграфах.
5.3. Взаимодействие биологических систем.
Математическая теория игр.
Как видим, главной особенностью живых организмов, как управляемых систем, заключается в том, что они могут устойчиво сохранять накопленный в них порядок, только постоянно увеличивая свою популяцию. А это значит, что они обязательно должны вступать в конфликт между собой или между различными популяциями. По сути дела, жизнедеятельность следует рассматривать как постоянный конфликт между различными формами упорядоченных систем.
В настоящее время в кибернетике активно разрабатывается математическая теория игр [18]. Смысл этой теории в том, чтобы понять, кто из двух (трёх, четырёх,...) конфликтующих сторон добьётся выигрыша (применительно к конфликту биологических систем – навяжет свой порядок).
Основными элементами теории игр являются:
Конфликтующие субъекты (игроки, соперники, партнёры) – управляемые системы, каждая из которых стремится навязать природе порядок, отличающийся от порядка соперников.
Природа (окружающая среда, шахматная доска с фигурами, колода карт, рынок и т.п.) – пассивный объект воздействия субъектов игры, меняющий свою конфигурацию (порядок) после каждого хода субъектов или независимо от них.
Ходы игроков и природы – единичные (простейшие) изменения природы под воздействием субъектов игры (личные ходы) или спонтанно в результате произвольного изменения самой природы (случайные ходы).
Правила игры – перечень всех возможных вариантов ходов, а также конечная конфигурация природы, определяемая как победа одной из конфликтующих сторон и поражение другой. В правила включается также процедура выполнения ходов партнёрами: поочерёдная, в зависимости от результатов предыдущего хода, от сложившейся конфигурации природы и т.п.
Ситуация (позиция, вариант) – конфигурация природы, сложившаяся спустя несколько личных и случайных ходов.
Выбор – конкретный ход, который выбирает один из конфликтующих субъектов из всего множества разрешённых для него ходов; исход – конкретный ход, осуществляемый природой.
Лучше всего
понять основные количественные
соотношения теории игр можно на примере
игры[18], включающей в себя двух игроков
и природу. Порядок выполнения ходов
следующий: 1
игрок – природа –
II
игрок –
природа – 1
игрок –
природа... и. т. д. В этом случае игра
делится на циклы, каждый из которых
состоит из одного хода обоих игроков и
двух ходов природы. После каждого
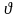 - ого цикла
у игрока 1
появляется возможность сделать n1J
вариантов ходов, у игрока II
– n2J
вариантов. У природы число ходов в два
раза больше, зато число возможных
вариантов nn
не меняется. Например, если в качестве
природы выступает игральная кость, то
число возможных вариантов всегда равно
шести. В шахматах или шашках ходы природы
вообще отсутствуют (число вариантов
равно нулю).
- ого цикла
у игрока 1
появляется возможность сделать n1J
вариантов ходов, у игрока II
– n2J
вариантов. У природы число ходов в два
раза больше, зато число возможных
вариантов nn
не меняется. Например, если в качестве
природы выступает игральная кость, то
число возможных вариантов всегда равно
шести. В шахматах или шашках ходы природы
вообще отсутствуют (число вариантов
равно нулю).
Игра заканчивается тогда, когда у кого-то из игроков число возможных вариантов станет равно нулю (т.е. либо n1Jmax = 0, либо n2Jmax = 0). Эта ситуация воспринимается как победа противоположной стороны, лишившей данного игрока возможности к дальнейшим действиям (ходам).
Главным понятием игры является стратегия – последовательность выборов одного игрока в ответ на предшествующие ходы соперника и исходы природы от первого до последнего хода. Вариантов таких последовательностей в каждой игре, даже если она относительно проста, чрезвычайно много. Однако ясно, что одна часть множества стратегий приведёт первого игрока к победе, а второго к поражению, а другая – к противоположной ситуации.
Приведём простейший пример: игру в «крестики-нолики» – рисунки 5.2 и 5.3. Как известно, цель каждого игрока – разместить три принадлежащие ему фигуры – крестики игроком I и нолики игроком II – так, чтобы их можно было соединить прямой линией, т.е. либо занять ими три клетки горизонтальной линии, либо три вертикальной, либо три диагональной линии. Чтобы избежать ничьей, иногда к этому условию добавляют ещё одно – в случае, если ни одному из игроков не удаётся выстроить три свои фигуры по прямой линии, выигрывает тот, кто делает последний ход. В этом случае подразделяют «чистый» выигрыш (аналог нокаута в боксе) и выигрыш «по очкам». Нетрудно заключить, что «по очкам» всегда выигрывает (при отсутствии третьего партнёра – природы) игрок I. Это вынуждает игрока II выбирать наиболее агрессивную стратегию.
Ясно, что игрок I, начинающий игру, может сделать девять различных ходов. Игрок II может ответить 8 ходами, так как первая клетка уже занята. В следующий цикл у игрока I остаётся всего семь вариантов ходов. Далее у игрока II остаётся 6 ходов. В следующем цикле у игрока I есть 5 ходов, однако их число фактически меньше, так как при некоторой конфигурации крестиков и ноликов, поставленных в предшествующих циклах, этот игрок выигрывает, поставив три крестика по прямой линии.
На рисунке 5.2, а показаны некоторые варианты выигрыша игрока I на третьем ходу. На самом деле их больше. Другие варианты можно получить из изображённых на рисунке 5.2, а путём симметричного отображения относительно горизонтальной, двух диагональных или вертикальной средней линии. На рисунке возле каждого квадрата показано число симметричных вариантов, соответствующих изображенной на нём конфигурации крестиков и ноликов. Как видим, у игрока I всего 120 стратегий, позволяющих выиграть на третьем ходу (из 9!/4!3!2!=1260 вариантов – множество из девяти клеток подразделяется на три подмножества – крестиков(3 клетки), ноликов (2 клетки) и пустых (4 клетки)).
Согласно §1.6 и [15 ] его статистический вес Р = 9!/4!3!2!. Нетрудно заключить, что 1140 положений на квадрате позволяют игроку II продолжить игру. Третий ход игрока II приводит к 9!/3!3!3! – 120= 1560 вариантам. Часть из них означает его победу. На рисунке 5.2, б так же, как на рисунке 5.2, а, показаны базовые варианты победы игрока II на третьем ходу. Возле каждого квадрата по-прежнему изображаем число симметричных конфигураций, соответствующих представленной на рисунке. Как видим, у игрока II на третьем ходу 140 вариантов выигрыша. Оставшиеся 1560 – 140 = 1420 конфигураций позволяют игроку I продолжить игру, делая четвёртый ход. На рисунке 5.2, в даны базовые конфигурации выигрыша этого игрока на этом ходу и число соответствующих каждой из них симметрий. Как видим, общее число выигранных игроком I на четвёртом ходу позиций равно 256, а оставшихся 9!/4!3!2! – 120 – 140 – 256 = 744. Эти оставшиеся позиции позволяют игроку II сделать четвёртый, последний ход,96 вариантов которого приносят ему выигрыш – см. рисунок 5.2, г.
У игрока I остаётся ещё 9!/4!4! – 96 –256 – 140 – 120 = 18 позиций, чтобы закончить игру и выиграть партию. (На самом деле у игрока I остаётся всего 9 ходов, так как каждым двум разным позициям соответствует одна свободная клетка). Таким образом, у игрока I всего 120 + 256 + 9 = 385 выигрышных стратегий, а у игрока II – 140 + 96 = 236 стратегий, т.е. в 1,63 раза меньше.
Следует отметить, что все проведённые расчёты исходят из того, что каждый из игроков делает случайные ходы и не «задумывается» об их последствиях – не просчитывает варианты вперёд. Тем не менее, даже в этом случае порядок на доске – в «природе» – увеличивается на
 Рисунок
5.2.Варианты выигрыша на третьем ходу
игры в крестики
– нолики
Рисунок
5.2.Варианты выигрыша на третьем ходу
игры в крестики
– нолики
DI = log2(385+236) = 9.28 бит,
где величина, стоящая под знаком логарифма – количество всех возможных вариантов до начала игры. Ясно, что стремление «выиграть» ведёт к увеличению степени интеллектуальности игрока, которая определяется способностью выбрать оптимальную стратегию, т.е. как можно лучше просчитать варианты развития событий.
Проанализируем, как можно выбрать оптимальную стратегию игры для игроков I и II. Во-первых, простой анализ показывает, что у игрока I не 9, а только три независимых друг от друга варианта первого хода – рисунок 5.3, а, так как остальные шесть ходов – зеркальные отображения одного из приведённых
Во-вторых, приведённые на рисунке 5.3, а варианты не эквивалентны друг другу: первый (крестик в центре доски) ведёт к четырём выигрышным стратегиям (на рисунке они обозначены стрелками, соединяющими клетки, занятие которых крестиками соответствует «чистой» победе); второй – к двум, третий – к трём. Ясно, что игроку I разумнее всего расположить крестик в центре доски. В ответ на этот ход у игрока II всего два варианта независимых хода – рисунок 5.3, б. Первый обеспечивает две победные стратегии – см. рисунок 5.3, б, а второй – только одну. Ясно, что оптимален для игрока II первый вариант.
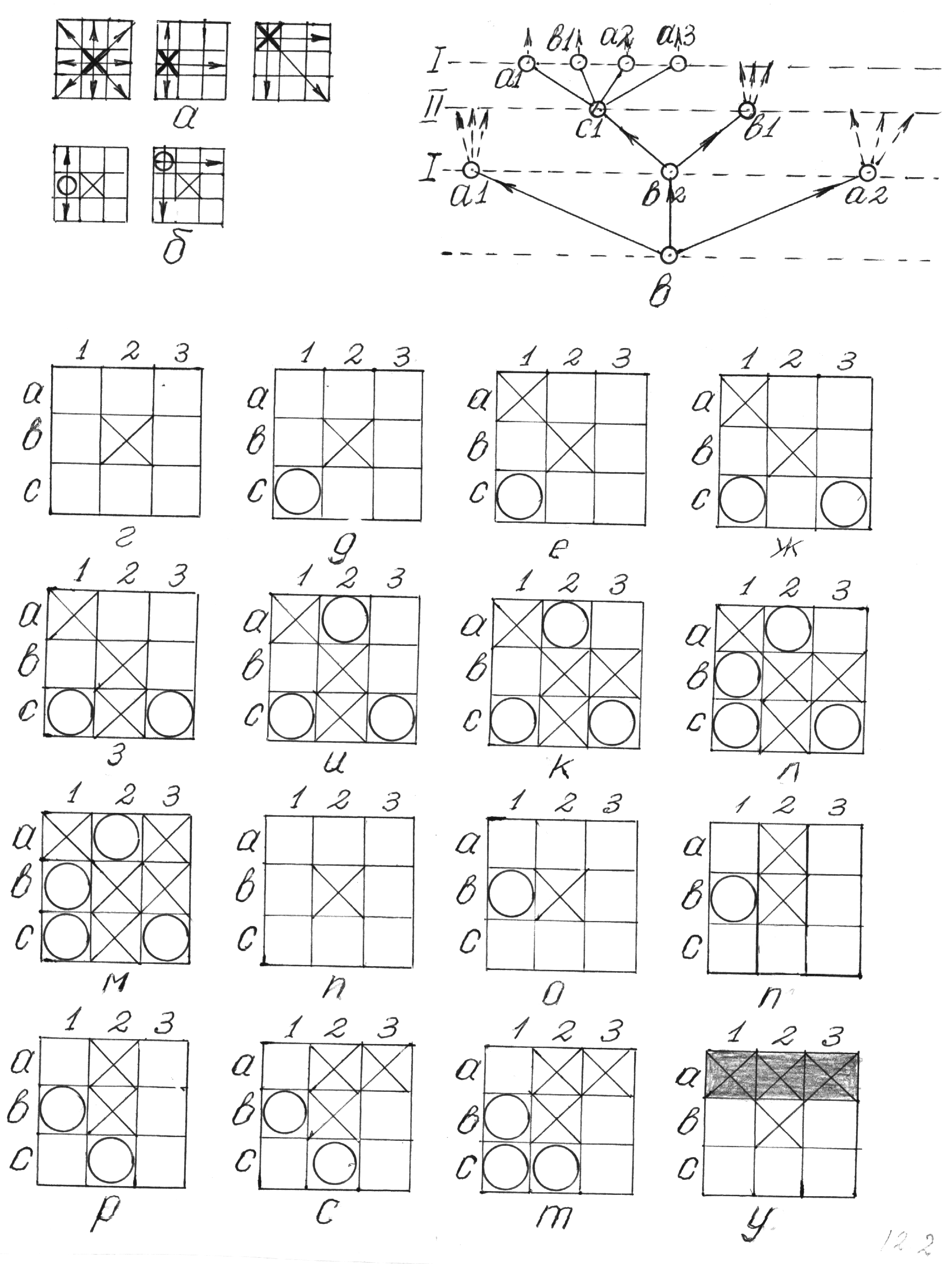 Рисунок 5.3
Рисунок 5.3
Оптимизация игры в «крестики-нолики»
И действительно, он, в конечном счёте, приводит к победе игрока I «по очкам» - рисунок 5.3, г-м, в то время как второй вариант – к «чистой» победе – рисунок 5., н-у. Стратегии игры можно изобразить графом – рисунок 5.3, в (см. ниже).
Основываясь на этом примере, изложим общие положения теории игр. Множество стратегий игрока I обозначим Х, а игрока II – Y, а их конкретные стратегии – х, у. Кроме этих стратегий существует стратегия случайных ходов природы h. Например, в указанной игре можно добавить ещё одно условие: после поочередного хода обоих игроков подбрасывается монета, и если она выпадает цифрой, то игрок II дополнительно ставит по своему усмотрению крестик (т.е. как бы подыгрывает игроку I). Если монета падает вверх гербом, то, наоборот, игрок I ставит нолик.
Совокупность стратегий h образует третье множество Н. Конкретная партия g представляет совокупность стратегий
g = g{x, y, h}.
Результат этой партии можно обозначить числом L, равным 1, если выигрывает игрок I, или 0, если выигрывает игрок II. Ясно, что L есть функция стратегий x, y, h:
L = L (x, y, h).
Она называется [15] функцией потерь. Учитывая, что стратегия h – случайная, можно определить её вероятность р(h). Например, при подбрасывании монеты р(h) = 0.5.
Тогда можно определить среднее значение потерь:
L(x,y) = ΣL(x,y,h)p(h); h Î H. (5.2)
В этом случае L(x,y) принимает любые значения от 0 до 1. Таким образом, математически игра G определяется тройкой множеств
G = {X, Y, L}, (5.3)
где Х, Y – некоторые множества, а L – числовая функция, определённая на произведении Х´Y. Элементы множеств хiÎ X, yjÎ Y называются стратегиями игроков I и II, а функция L – функцией потерь. Числа m и n определяют число стратегий первого и второго игроков.
Цель игры для первого игрока – максимизировать L, а второго – её минимизировать. Всего значений L – m´n и они образуют матрицу:
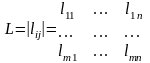 .
(5.4)
.
(5.4)
Особую роль играют так называемые статистические игры. Они отличаются от предыдущих – стратегических игр – тем, что в них всего один игрок и природа. Игрок в статистических играх именуется статистиком. Ясно, что в этих играх функция потерь зависит от всевозможных состояний природы h и стратегий статистика х:
L = L (x, h). (5.5)
Для того чтобы представить себе взаимодействие стратегий в процессе игры, обычно прибегают к древовидному графу. На рисунке 5.3, в построен граф игры в крестики-нолики, описанной выше без «статистического» игрока.
Как видим, игры, независимо от того, где они происходят, носят очевидный антиэнтропийный характер: победу в них в подавляющем большинстве случаев одерживает тот партнёр, у которого степень упорядоченности (точнее степень устойчивости упорядоченности) выше. Результаты игры приводят к повышению меры упорядоченности в природе (следует не забывать, что под «природой» в теории игр понимается статистический партнёр, т.е. партнёр, действия которого носят случайный, сумбурный характер).
Напомним, что конфликтные ситуации («игры») возникли на Земле в результате появления биологических систем.