

3.16.2. Долговременная память
Классификация видов памяти. И. С. Бериташвили (1975) различал образную, условнорефлекторную, эмоциональную и словесно—логическую память. Под образной памятью человека и животных понимают сохранение в памяти и репродукцию однажды воспринятого жизненно важного объекта; под эмоциональной памятью — воспроизведение пережитого ранее эмоционального состояния при повторном воздействии раздражителей, обусловивших первичное переживание этого состояния.
Эмоциональная память обладает следующими особенностями: 1) она надмодальна, т. е. ее формирование и воспроизведение может происходить при любых сенсорных воздействиях; 2) она реализуется очень быстро и часто с первого раза; 3) она характеризуется непроизвольностью запоминания и воспроизведения информации, т. е. обеспечивает пополнение подсознательной сферы человеческой психики.
Условнорефлекторная память проявляется в виде воспроизведения условных двигательных и секреторных реакций или заученных привычных движений спустя длительное время после их образования.
Наконец, словесно—логическая память — это память на словесные сигналы, обозначающие внешние объекты, события, внутренние переживания и свои собственные действия.
В любом случае происходит фиксация образов, предметов и явлений, включающая в свой состав по крайней мере три этапа: формирование энграммы, сортировку и выделение новой информации, долговременное хранение значимой для организации информации.
Основная гипотеза базируется на признании того, что фиксация следа памяти связана со стойкими изменениями синаптической проводимости в пределах определенного нейронного ансамбля.
В основе долговременной памяти лежат сложные структурно—химические преобразования на системном и клеточном уровнях головного мозга.
Под влиянием научения увеличивается количество холинорецепторов, что может быть следствием либо синтеза новых рецепторных молекул, либо демаскировки и активации уже существующих рецепторных белков. Установлено повышение чувствительности кортикальных нейронов к ацетилхолину в процессе образования условного рефлекса. Активация рецепторов постсинаптической мембраны облегчает научение, ускоряет фиксацию, способствует извлечению следа из памяти. И наоборот, антагонисты ацетилхолина нарушают научение и воспроизведение, вызывая амнезию (потерю памяти).
Катехоламинергическая и серотонинергическая системы принимают самое непосредственное участие в механизмах долговременной памяти. Установлено, что научение животных в моделях с электрокожным подкреплением условных рефлексов сопровождается активацией адренергических систем мозга, а научение с пищевым подкреплением — снижением метаболизма и уровня норадреналина в головном мозгу. '
Предполагают, что норадреналин пролонгирует активность нейронов, вызванную предъявлением условного стимула, и этим облегчает формирование условного рефлекса. Серотонинергической системе принадлежит важная роль в процессах консолидации следов памяти. Серотонин ускоряет научение и удлиняет сохранение навыков, выработанных на эмоционально положительном подкреплении, нарушая выполнение и сохранение защитно—оборонительных реакций. Согласно существующей концепции, моноамины участвуют в процессах научения и памяти опосредованно, через нейрохимическое обеспечение положительных и отрицательных эмоций. Серотонин причастен к формированию эмоционально положительных, а норадреналин — эмоционально отрицательных состояний.
Существует гипотеза, согласно которой холинергические механизмы мозга обеспечивают информационную составляющую процессов научения. Роль моноаминергических систем мозга сводится к облегчению подкрепляющих и эмоционально—мотивационных составляющих процесса научения и памяти, причем холинергическая система находится под модулирующим влиянием моноаминергической системы. Стимуляция моноаминергических механизмов перестраивает хемореактивные свойства, которые при определенных условиях могут закрепляться, обеспечивая создание и сохранение многонейронной констелляции — энграммы. Если адренергические механизмы мозга в большей мере причастны к формированию временных связей, то серотонинергические — к их фиксации.
Процессы долговременной памяти зависят также от уровня метаболизма ГАМК, глутаминовой кислоты, вторичных посредников (циклических нуклеотидов и ионов кальция).
Участие информационных макромолекул (нуклеиновых кислот и белков) в ключевых процессах научения и памяти не вызывает сомнений. Согласно одной группе гипотез, научение и память связаны с кодированием приобретенных форм поведения в информационных макромолекулах. Согласно другой группе гипотез, которая исходит из взаимосвязи генома и синтеза специфических белков нервной клетки, на основе функционального объединения нейронов возникает структурное их объединение, представляющее собой энграмму памяти.
Первая группа гипотез базируется на аргументах о качественном изменении РНК и белков при научении и возможности переноса памяти от обученного мозга к необученному с помощью РНК или полипептидов. Установлено, что для процессов консолидации и формирования долговременной памяти необходим полноценный белковый синтез. В формировании устойчивости созданной энграммы должна существовать система обновления специфических рецепторных белков либо должны возникать стабильные модификации ДНК, в
-
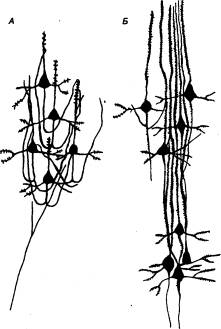
Рис. 3.50 Основные типы нейронных объединений в лобной коре обезьян
А — нейронная «ловушка»; Б — вертикальный модуль.
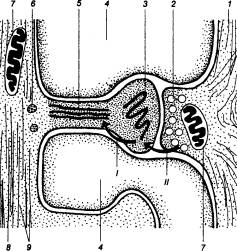
Рис. 3.51 Аксошипиковый транзиторный синапс
1 — аксон, 2 — синаптические везикулы, 3 — шипиковый аппарат, 4 — нейроглия, 5 — молекула актина, б — рибосомы, 7 — митохондрия, 8 — дендрит, 9 — дендритные трубочки; I — влияние пресинаптического возбуждения на постсинаптическую мембрану, вызывающее в головке шипика возрастание концентрации Са2+; II — влияние Са2+ на молекулы актина, приводящее к сокращению ножки шипика и облегченному проведению местного потенциала к стволу дендрита.
результате которых в нейронах образуется и поддерживается пожизненно синтез любого нейроспецифического белка или, наоборот, необратимо выключается синтез маскирующего белка.
Полагают, что повторная импульсация в нейроне сопровождается увеличением концентрации кальция в постсинаптической мембране. Это активирует фермент — кальций—зависимую протеиназу, которая расщепляет один из белков мембраны. Его расщепление высвобождает замаскированные ранее неактивные белковые глутамат—рецепторы. С возрастанием числа активных глутаматрецепторов возникает состояние повышенной проводимости аксошипикового синапса (в течение 3—6 сут).
Аксошипиковые контакты — наиболее пластичное соединение между нейронами, которое может быть ответственно за эффективность синаптической проводимости. Последнее достигается изменением диаметра самого шипика, что меняет, в свою очередь, сопротивление мембраны. Это обеспечивается наличием в ножке шипика контрактильного аппарата в форме молекул актомиозина. Их активация возникает при высвобождении ионов кальция из депо, коим является расположенный в головке шипиковый аппарат (рис. 3.50, 3.51). Высвобождение ионов кальция происходит при действии медиаторов на постсинаптическую мембрану. Сокращение молекул актомиозина приводит к укорочению и утолщению ножки шипика, вследствие чего меняется сопротивление и проведение электрического потенциала к дендритному стволу.
Для понимания механизмов «переноса памяти» целесообразно использовать гипотезу И. П. Ашмарина об участии иммунологических механизмов в долговременной памяти (рис. 3.52). Если представить, что после прохождения импульсов через синапс усиливается синтез специфических белков—антигенов, то их избыток должен
|
Рис. 3.52 Представление об иммунохимическом механизме формирования памяти (по Ашмарину) КБ — катионные белки, открывающие путь комплексам синаптического антигена с РНК через оболочку клетки; Л — клетка глии (аналог лимфоцитов); М — клетка глии (аналог макрофагов); 1, 2, ? — участки синаптического антигена (зона синтеза заштрихована); 4 — комплекс фрагмента синаптического антигена с РНК; 5 — антитела для синапсов. Тонкими линиями изображены мРНК, толстыми — ДНК хромосом,. |
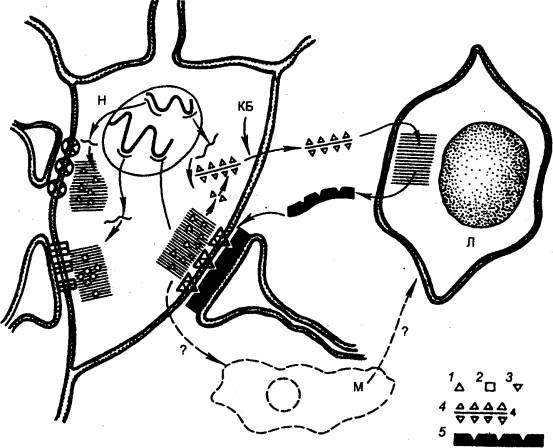
выходить в околосинаптическое пространство. Эти белки взаимодействуют с рядом расположенными клонами клеток астроцитарной глии, индуцируют их размножение и образование антител. Последние специфически взаимодействуют с постсинаптическими мембранами тех же нейронов и облегчают проводимость в соответствующих синапсах. Данный клон астроцитов сохраняется в течение жизни. Следовательно, действующим началом «переноса памяти» может быть избыточный антиген пептидной природы, который способен автоматически найти в мозгу реципиента либо соответствующую клетку глии либо синапс. '
Специальные исследования на животных с генетическим дефицитом вазопрессина и нарушением памяти обнаружили ее улучшение после инъекции дополнительных количеств этого гормона. Противоположное действие — нарушение сохранения выработанных навыков — оказывает другой гормон гипофиза — окситоцин. Эндорфины и энкефалины (эндогенные опиаты) оказывают выраженное влияние на научение и память — замедляют угашение
условных рефлексов, улучшают их сохранение, хотя и ухудшают их формирование.
Нейропептиды обнаружены в аксонных окончаниях нейронов вместе с классическими медиаторами. Нейропептиды могут усилить или ослабить действие медиатора. Нейропептиды—спутники вместе с медиаторами создают и поддерживают на постсинаптической мембране специфические рецепторные мозаичные наборы, способствующие быстрому проведению определенного вида возбуждения. Пептид—спутник повышает сродство рецептора к основному медиатору, он более стабилен, чем основной медиатор, что обеспечивает пролонгированное облегчение проведения через синапс.
Вероятно, все формы нейрологической памяти обеспечиваются системой межнейронных взаимодействий, участие же информационных молекул проявляется лишь во включении или выключении различных участков предсуществующего генома, а не в синтезе новых нуклеотидных или аминокислотных последовательностей.