

8. Клеточная стенка грамположительных бактерий.

Фиолетовые\синии палочки)
Суть метода Грама в том, что исследуемую среду обрабатывают
анилиновым красителем, фиксируют йодом, а затем смывают спиртом.
При этом грамотрицательные бактерии обесцвечиваются, а
грамположительные приобретают синюю окраску. После повторной
обработки (обычно сафранин), который окрашивает все
грамотрицательные бактерии в красный или розовый цвет, причем
погибшие микроорганизмы окрашиваются ярче.
Это происходит из-за наличия внешней мембраны, препятствующей
проникновению красителя внутрь клетки. Сам по себе тест полезен при
классификации бактерий и разделении их на две группы относительно
строения их клеточной стенки. Из-за своей более мощной и
непроницаемой клеточной стенки грамотрицательные бактерии более
устойчивы к антителам и антибактериальным препаратам. Поэтому
заболевания, вызываемые ими, очень сложно лечить, сложнее чем (+)
Метод Грама позволил разделить все микроорганизмы на две большие группы.
Их особенности и характеристики помогают выбрать более подходящее
лечение инфекционных заболеваний. Грамположительные бактерии, которые
быстро окрашиваются в синий цвет анилиновым красителем, образуют споры,
экзотоксины, поэтому довольно опасны для здоровья. Но их оболочка
проницаема для антибактериальных препаратов.
|
Признак и компоненты |
Грамположительные |
Грамотрицательные |
|
1. Структура |
однородная |
слоистая |
|
2. Толщина |
20 – 60нм |
14 – 18 нм |
|
3. Ригидный и пластичный |
связаны ковалентно |
связаны лабильно |
|
4. Пептидогликан |
до 90% сухой массы, предст. 5 – 6 слоями, часто не содержит диаминопимелиновую кислоту |
до 5 – 10% сухой массы предст. 1 – 2 слоями, содержит диаминопимелиновую кислоту |
|
5. Тейховые кислоты |
до 50% сухой массы |
отсутствуют |
|
6. Наружная мембрана |
у некоторых имеются токсические гликолипиды; липополисахариды отсутствуют |
представлена липопротеинами, фосфолипидами, липополисахаридами |
|
7. Белки |
белки, определяющие антигенную специфичность |
белки – порины |
|
8. Чувствительность к пенициллинам и лизоциму |
высоко чувствительна |
менее чувствительна |
|
Представители |
бациллы, клостридии, коринебактерии, стафилококки, стрептококки, пептококки |
энтеробактерии, вибрионы, нейссерии, вейлонеллы, бактероиды, спириллы, псевдомонады |
На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты делятся на две большие группы. Было обнаружено, что если фиксированные клетки бактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются (этот способ был впервые предложен в 1884 г. датским ученым X. Грамом (Ch.Gram), занимавшимся окрашиванием тканей. Позднее он был использован для бактерий).
У некоторых бактерий положительная реакция при окрашивании описанным способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
Клеточные стенки грамположительных и грамотрицательных прокариот резко различаются как по химическому составу (таблица 1), так и по ультраструктуре (рис. 1).
В состав клеточной стенки прокариот входят семь различных групп химических веществ, при этом пептидогликан (структурный полимер, являющийся основным опорным элементом клеточной стенки) присутствует только в клеточной стенке. У грамположительных бактерий он составляет основную массу вещества клеточной стенки (от 40 до 90 %), у грамотрицательных -- содержание пептидогликана значительно меньше (1 --10 %). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных бактерий, содержит от 20 до 50 % этого гетерополимера.
Под электронным микроскопом клеточная стенка грамположительных бактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных бактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2--3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8--10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных бактерий получил название наружной мембраны.
Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране (ЦПМ) в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп бактерий, характерно только для грамотрицательных форм (рис. 1).
Опорный скелет бактериальной стенки состоит из однородного полимера, обозначаемого как мукопептид, глюкозаминопептид, гликопептид, пептидогликана или муреин (лат. мurus - стенка). В настоящее время общеприняты два последних наименования.
Эта макромолекула - гетерополимер, построенный из цепочек, в которых чередуются остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты (N-ацетилглюкозаминлактата), соединенные между собой Я-1,4-гликозидными связями. Такие неразветвленные гетерополимерные цепи образуют основу муреина. Остатки мурамомвой кислоты через лактильные группы соединены пептидной связью с аминокислотами. К типичным аминокислотам муреина относятся L-аланин, D-глутаминовая кислота, мезо-диаминопимелиновая кислота или L-лизин и D-аланин. Диаминокислоты мезо- (или LL-) диаминопимелиновая кислота и L-лизин играют большую роль в межмолекулярных сшивках, так как образуют пептидные связи с участием обеих аминогрупп и, таким образом, могут связать две гетерополимерные цепи между собой (рис. 2). Место диаминопимелиновой кислоты или лизина могут занять орнитин или диаминомасляная кислота. Пептидными мостиками гетерополимерные цепи связаны между собой в мешкообразную гигантскую молекулу - муреиновый мешок.
Особого внимания заслуживает то, что в бактериальной стенке содержатся структуры и вещества, которых нет у животных и растений: таковы, например, чередующаяся последовательность N-ацетилглюкозамина и N-ацетилмурамовой кислоты; не встречающаяся в составе белков мезо-диаминопимелиновая кислота; D-формы аланина и глутаминовой кислоты. Эти структурные элементы составляют ахиллесову пяту бактерий, используемую врачами в борьбе с инфекцией. По компонентам и структуре клеточной стенки и биохимическим механизмам ее синтеза бактерии коренным образом отличаются от животных и растений. Поэтому лекарственные препараты, специфически воздействующие только на бактериальные стенки и на процесс их синтеза, должны быть безвредными для высших организмов.
Наличие в клеточных стенках пептидогликанового слоя характерная особенность всех прокариот. Исключение составляют только архебактерии и немногие другие группы и виды.

Клеточная стенка грамположительных бактерий.
У грамположительных бактерий доля муреиновой сетки составляет 30-70% сухой массы клеточной стенки (толщиной в 40 слоев). Вместо м-диаминопимелиновой кислоты часто содержатся LL-диаминопимелиновая кислота или лизин. У Staphylococcus aureus тетрапептидные боковые цепи мурамовой кислоты связаны между собой межпептидными (например, пентаглициновыми) цепочками. Участвующие в образовании таких структур аминокислоты варьируют от вида к виду. Видоспецифическое строение опорного каркаса представляет собой хороший таксономический признак. В клеточной стенке грамположительных бактерий полисахариды, если они вообще имеются, связаны между собой ковалентно. Содержание белков невелико. Характерная особенность - наличие тейхоевых кислот (рис. 3); это цепи, состоящие из 8-50 остатков глицерола или рибитола, связанных между собой фосфатными мостиками. Некоторые из тейхоевых кислот содержат эритритол или маннитол. Тейхоевые кислоты, вероятно, через фосфат связаны с муреином по типу амида.
Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений - тейхоевые кислоты , представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями (рис. 5). Тейхоевые кислоты, стоящие из остатков глицерина (глицеринтейхоевая) или рибита (рибиттейхоевая), соединенных между собой пирофосфатными мостиками. К доступным гидроксильным группам этих спиртов часто присоединены аминокислоты и моносахариды, которые во многом определяют биологические и иммунологические особенности заболеваний, вызываемых грамположительными бактериями. Некоторые тейхоевые кислоты (липотейхоевые кислоты) ковалентно связаны с гликолипидами мембран.
В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой.
Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией. Клеточная стенка бацилл, например Bacillus subtilis , приблизительно соответствует толщине 40 молекул пептидогликана. В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1-6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор.


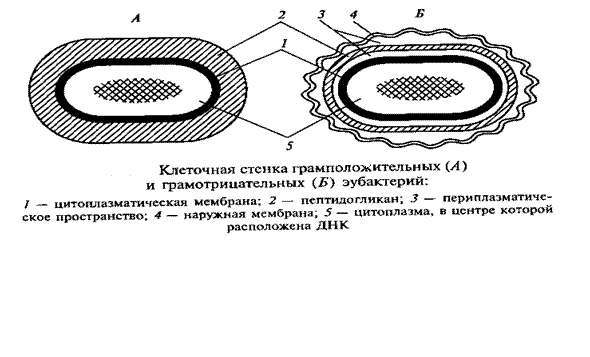
9. Клеточная стенка грамотрицательных бактерий. https://medlec.org/lek2-65262.html ДОП ПОЧИТАТЬ Клеточная стенка грамотрицательных бактерий. У грамотрицательных бактерий муреиновая сеть однослойная (рис. 4) и составляет менее 10% сухой массы клеточной стенки (у Escherichia coli). Муреин содержит только мезодиаминопимелиновую кислоту и не содержит лизина; межпептидные мостики отсутствуют. Строение муреинового мешка у всех грамотрицательных бактерий одинаково.
У грамотрицательных бактерий строение клеточной стенки намного сложнее, чем у грамположительных (рис. 4). В ее состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных бактерий содержание этого гетерополиме-ра колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки -- наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (рис. 4). Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30--40 % ее поверхности и локализованный во внешнем слое.
Белки
наружной мембраны можно разделить на
основные и минорные. Основные белки
представлены небольшим числом различных
видов, но составляют почти 80 % всех белков
наружной мембраны. Одна из функций этих
белков -- формирование в мембране
гидрофильных пор диаметром примерно 1
нм, через которые осуществляется
неспецифическая диффузия молекул с
массой до 600--900 Да (Да -- дальтон, или
единица атомной массы, равен 1,66033-1027
кг.). Это означает, что через такие поры
могут проходить сахара, аминокислоты,
небольшие олигосахариды и пептиды.
Белки, пронизывающие наружную мембрану
насквозь и образующие гидрофильные
поры, называют поринами (рис. 4). Минорные
белки наружной мембраны представлены
гораздо большим числом видов. Их основная
функция -- транспортная и рецепторная.
Примером минорных белков могут служить
белки, ответственные за специфический
транспорт в клетку железосодержащих
соединений.

Структура грамположительной клеточной стенки. ЦПМ - цитоплазматическая мембрана. ПГ - пептидогликан. ЛТК - липотейхоевые кислоты
Структура грамотрицательной клеточной стенки. ВМ - внешняя мембрана (содержит фосфолипиды на внутренней поверхности и липополисахариды - на внешней).ПГ - пептидогликан. ЦПМ - цитоплазматичеукая мембрана. ЛПС - липополисахарид, состоит из липидной части, которая обращена внутрь внешней мембраны и формирует ее гидрофобную область, и полисахаридной части, которая обращена во внешнюю среду. ЛП -липопротеиды.
Помимо слоев клеточной стенки, типичных для большинства грамотрицательных бактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной клеточной мембраны. Однако до настоящего времени не ясно, относятся ли они к клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла.
Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, что говорит об эластичности их клеточной стенки и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных бактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий -- чрезвычайно низкая сшитость ее пептидогликанового компонента.

Рисунок 1 (-) слева
Грамотрицателъные бактерии имеют сравнительно тонкую клеточную стенку. В ней выделяют два слоя — пластичный и ригидный. Последний образован одним, редко двумя слоями пептидогликана, содержание которого составляет не более 20% сухой массы клеточной стенки. На пептидогликановом каркасе расположены фосфолипиды, липополисахариды (ЛПС) и белки, образующие пластичный слой. Толщина пластичного слоя значительно превышает размеры монослоя пептидогликана. Его компоненты расположены мозаично и могут образовывать дополнительную внешнюю мембрану либо переходить в капсулу. • Фосфолипиды клеточной стенки пластичного слоя прикреплены к пептидогликану липопротеинами, пересекающими периплазматическое пространство. Обработка детергентами (например, додецилсульфатом натрия) приводит к нарушению этих связей. Основное отличие внешнего фосфолипидного слоя от внутреннего ригидного — высокое содержание липополисахариды. • Липополисахариды клеточной стенки состоят из липидной части (липид А), базисной части молекулы полисахарида (сердцевина) и боковых полисахаридных цепей (рис. 4-5). Иммуногенные свойства проявляют боковые полисахаридные цепи и сердцевина. Боковые полисахаридные цепи отвечают за антигенную специфичность молекулы липополисахаридов и называются О-Аг. Липидная часть термоустойчива и отвечает за биологические эффекты ЛПС. Структура ЛПС имеет большое диагностическое значение, поскольку разные виды или серовары патогенных грамотрицательных бактерий отличаются друг от друга составом боковых цепей ЛПС внешней мембраны. • Белки, входящие в состав пластичного слоя, подразделяют (в зависимости от выполняемых функций) на основные (мажорные) и второстепенные (минорные). К мажорным белкам относят порины, образующие трансмембранные каналы, вовлечённые в транспорт ионов и гидрофильных соединений из внешней среды в периплазму. Минорные белки также могут участвовать в транспорте веществ через пластичный слой (путём облегчённой диффузии или активного транспорта молекул). Некоторые белки играют роль рецепторов для вирусов бактерий и бактериоцинов, а также для донорских пилей при конъюгации. Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий. Внешняя мембрана не пропускает молекулы с большой молекулярной массой, что можно рассматривать как фактор неспецифической устойчивости бактерий к некоторым антимикробным препаратам.
Источник: https://meduniver.com/Medical/Microbiology/43.html MedUniver
