

18. Поверхностные нежгутиковые структуры прокариот.
19. Жгутики. Расположение и функции.
Жгутик – это поверхностная структура бактериальной клетки, которая служит им для движения в жидких средах.
В
зависимости от
расположения
жгутиков, бактерии делятся на

1.Полюсные жгутики – один или более жгутиков расположены на одном (монополярно) или обоих (биполярно) полюсах клетки и основание параллельно длинной оси клетки. 2.Подполюсные жгутики (субполярные) – один или более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух ее концах. В основании – прямой угол с длинной осью клетки. 3.Боковые жгутики (латеральные) - один или более жгутиков в виде пучка расположены в средней точке одной из половин клетки. 4.Перитрихиальные жгутики – расположены по всей поверхности клетки по одному или пучками, полюса обычно их лишены. 5.Смешанные жгутики – два или несколько жгутиков расположены в разных точках клетки.
В зависимости от числа жгутиков, различают: 1) Атрихи – это бактерии без жгутиков. 2) Монотрихи- один полярно расположенный жгутик(как у холерного вибриона). У них есть всего один жгутик, при помощи которого они передвигаются 3) Лофотрихи-пучок жгутиков на одном конце. Для таких бактерий характерно расположение жгутиков только в одной части бактерии 4) Амфитрихи – пучки жгутиков с двух концов. 5) Перитрихи- множество жгутиков вокруг клетки (как у кишечной палочки). это бактерии, жгутики у которых располагаются равномерно по всей длине бактерии. Они позволяют передвигаться бактерии плавно, без рывков Функции: Эти структурные элементы клетки определяют ее подвижность. Чаще всего это тонкие нити, которые берут свое начало еще от цитоплазматической мембраны. Некоторые виды микробов имеют существенно больший жгутик, чем сама клетка-хозяин. Отростки способны проталкивать клетку в жидкой среде. Строение жгутика таково, что он может быстро перемещать тело-клетку, и при этом она будет преодолевать сравнительно большие расстояния. Движения эти совершаются по принципу пропеллера. Чтобы перемещаться, микробы используют один или несколько отростков. У некоторых микробов отростки могут быть дополнительным фактором патогенности (болезнетворности). Это можно объяснить с тем, что он способствует приближению патогенного микроорганизма к здоровой клетке. https://vseobiology.ru/mikrobiologiya/1766-11-zhgutiki-fimbrii-bakterij-stroenie-funktsii-mekhanizm-dvizheniya фимбрии и пили
-
20. Строение жгутика у грамположительных и грамотрицательных бактерий. Синтез жгутика.
-
Филамент (фибрилла, пропеллер) — полая белковая нить толщиной 10—20 нм и длиной 3—15 мкм, состоящая из флагеллина, субъединицы которого уложены по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от плазматической мембраны. По полости к собираемому в настоящий момент участку переносятся субъединицы флагеллина. Филамент жгутика – это относительно жесткая белковая спираль, закрученная против часовой стрелки с образованием центрального полого канала диаметром до 3 нм. Такая конструкция способствует формированию спиральной траектории движения нити.
-
Крюк (колено) — более толстое, чем филамент (20—45 нм), белковое (не флагеллиновое) образование. Крюк соединяет нить с базальным телом жгутика и состоит из двух типов белка: FlgE и FlgKl. Длина сочленения постоянна и составляет около 50 нм. Из-за изогнутой формы крюка при вращении мотора основание фибриллы описывает круг, благодаря чему возможно спиралевидное движение жгутика
-
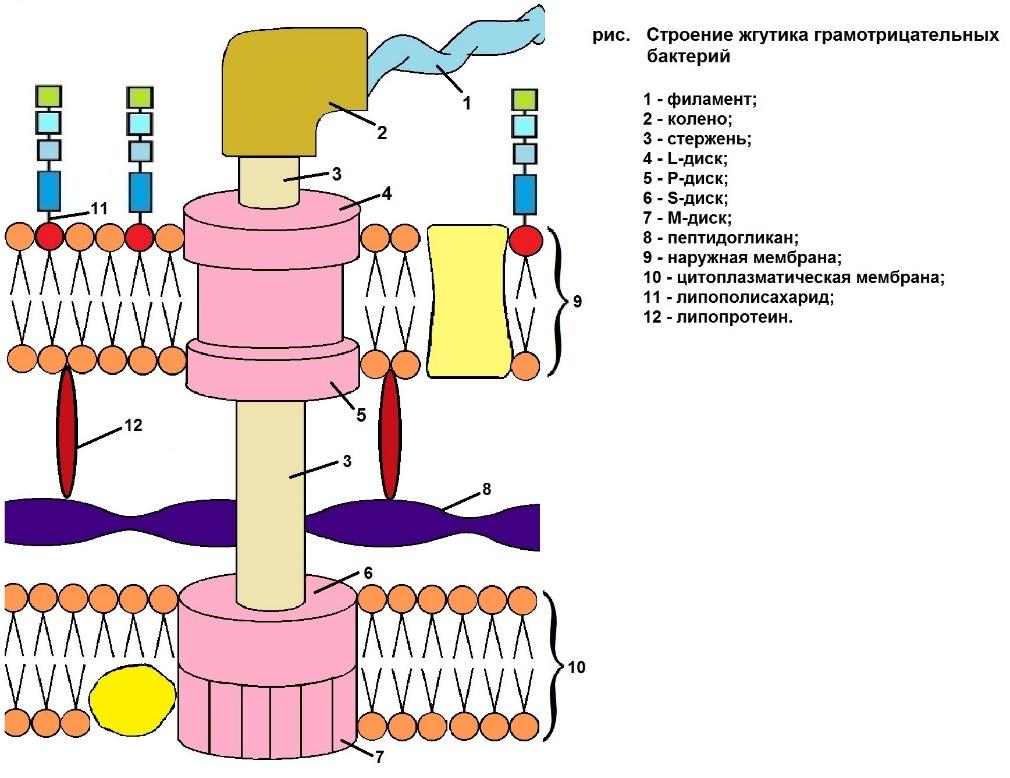
Базальное тело (трансмембранный мотор). Базальное тельце закреплено в клеточной стенке и цитоплазматической мембране бактерий. Эта субструктура выполняет не только фиксирующую функцию, но и является мотором жгутика. Строение и локализация базального тельца зависят от типа клеточной стенки микроорганизма. У грамотрицательных бактерий оно состоит из двух внутренних (M и S) и двух внешних (P и L) колец, нанизанных на соединенный с крюком стержень. В состав базального тела также входит экспортная система, транспортирующая белковые элементы для сборки жгутика.
Базальное тело представляет собой систему колец, находящихся в плазматической мембране и клеточной стенке бактерий. Два внутренних кольца — M и S-кольца (сейчас чаще рассматриваются как единое MS-кольцо) — являются обязательными элементами, причём M-кольцо находится в ЦПМ, а S — в периплазме грамотрицательных и пептидогликановом слое грамположительных бактерий. Ещё два кольца — P и L — есть только у грамотрицательных бактерий, они расположены в пептидогликановом слое и наружной мембране соответственно, неподвижны и лишь направляют стержень ротора мотора. Вокруг MS-кольца расположены статоры — белковые комплексы MotA4/MotB4, представляющие собой протонный канал (их может быть от 8 до 16).
Точный механизм работы базального тела неизвестен. Большинство исследователей полагает, что поступление протона из периплазмы или внешней среды в MotA4/MotB4 комплекс вызывает конформационные изменения белков, благодаря электростатическому взаимодействию или прямому контакту это изменение приводит к повороту MS-кольца, а его дальнейшее движение возвращает исходную конформацию комплексу и выталкивает протон в цитозоль.
Жгутик состоит из трех компонентов – спиральной жгутиковой нити (филамента) постоянной толщины, крючка (колена) и базального тельца. Крючок, к которому присоединена жгутиковая нить, имеет длину 30-45 нм и состоит из отличающегося от флагеллина белка. Он соединен с базальным тельцем, которое располагается в оболочке (в клеточной стенке и ЦПМ). Нити жгутиков приводит в движение мембранный шарнироподобный базальный крюк. Базальное тельце состоит из центрального стержня, заключенного в систему особых колец. Кольца выполняют роль «приводного диска» и «подшипника» на внутренней поверхности пептидогликанового слоя. Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор).
Вращение жгутика в клеточной стенке происходит из-за вращательного движения колец S и М относительно друг друга и обеспечивается за счет энергии трансмембранного градиента ионов водорода или натрия. Благодаря такому вращению происходит движение бактерий в наиболее благоприятном для них направлении. Жгутиковый аппарат обладает особым бинарным переключателем, который позволяет менять направление вращения жгутиков против часовой стрелки на противоположное. Таким образом, бактерии, получив химический сигнал из окружающей среды, изменяют направление движения и выбирают оптимальные условия обитания. Расположение кольцевых субъединиц в клеточной оболочке соответствует следующей схеме: "М" – цитоплазматическая мембрана; "S" – периплазматическое пространство или клеточная стенка у Г+-бактерий; "P" – пептидогликановый слой; "L" – наружняя мембрана. Внешние кольца P и L неподвижны и выполняют поддерживающую функцию. У грамположительных бактерий они отсутствуют.
У грамотрицательных бактерий две пары колец: внешняя (кольца L и Р) и внутренняя (кольца S и М). Кольца L и Р расположены внутри клеточной стенки (кольцо L в ЛПС, а кольцо Р – в слое пептидогликана). Они выполняют, очевидно, роль втулки для стержня. Внутренняя пара (кольца S и М) фиксирована на ЦПМ, причем кольцо S располагается в периплазматическом пространстве, а кольцо М – на ЦПМ или в ней. Жгутики грамположительных бактерий, имеющих более толстую гомогенную клеточную стенку, содержат только одну пару колец – S и М.

Вращательный тип движения присущ извитым формам, например, спирохетам. Они передвигаются за счет колебания собственного тела, очень подвижны. Жгутик работает как винт или пропеллер.
Синтез жгутика

Процесс синтеза жгутика эубактерий запускается экспрессией гена сtrA. Продуктом этого гена является белок CtrA. Синтез CtrA происходит сразу после перехода клетки из G0-фазы( период клеточного цикла, в течение которого клетки находятся в состоянии покоя и не делятся.) в S-фазу (в которой происходит репликация ДНК. Стадия интерфазы,). Обычно участок ДНК, содержащий ген сtrA, метилирован. Синтезу белка CtrA предшествует деметилирование ДНК, которая затем реплицируется. После этого происходит синтез CtrA и его фосфорилирование киназами. Ген сtrA имеет два промотора: Р1 и Р2. С первого промотора осуществляется ингибирование синтеза CtrA. Со второго промотора осуществляется стимулирование синтеза CtrA. Следует отметить, что белок CtrA найден не у всех эубактерий, и механизм синтеза жгутика не универсален.